

Microcircuitry for two types of achromatic ganglion cell in primate fovea
- PMID: 17344402
- PMCID: PMC6672494
- DOI: 10.1523/JNEUROSCI.4739-06.2007
Microcircuitry for two types of achromatic ganglion cell in primate fovea
Abstract
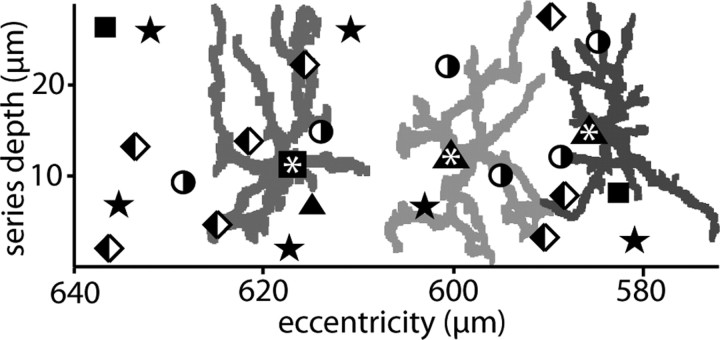
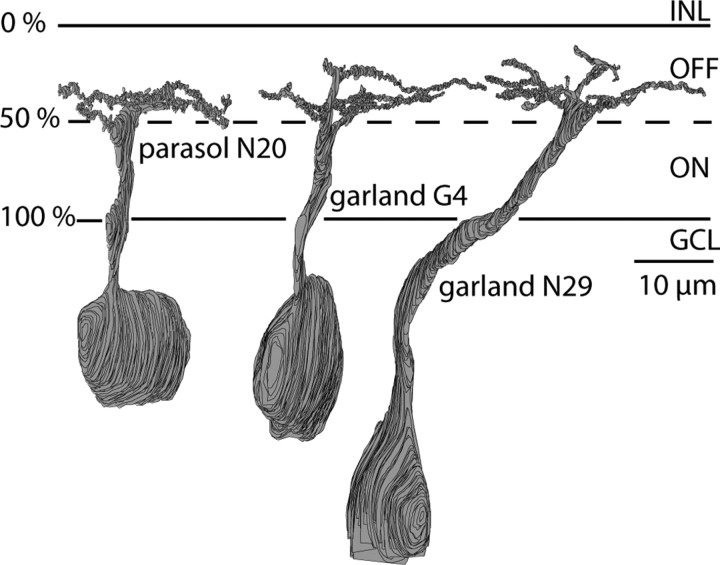
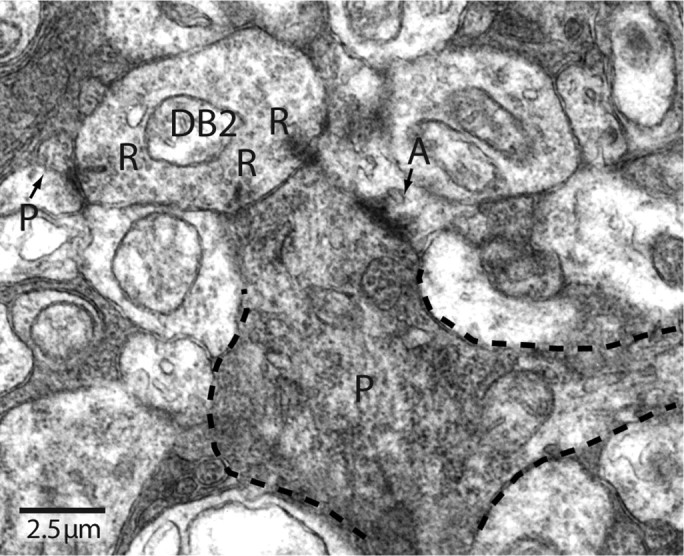
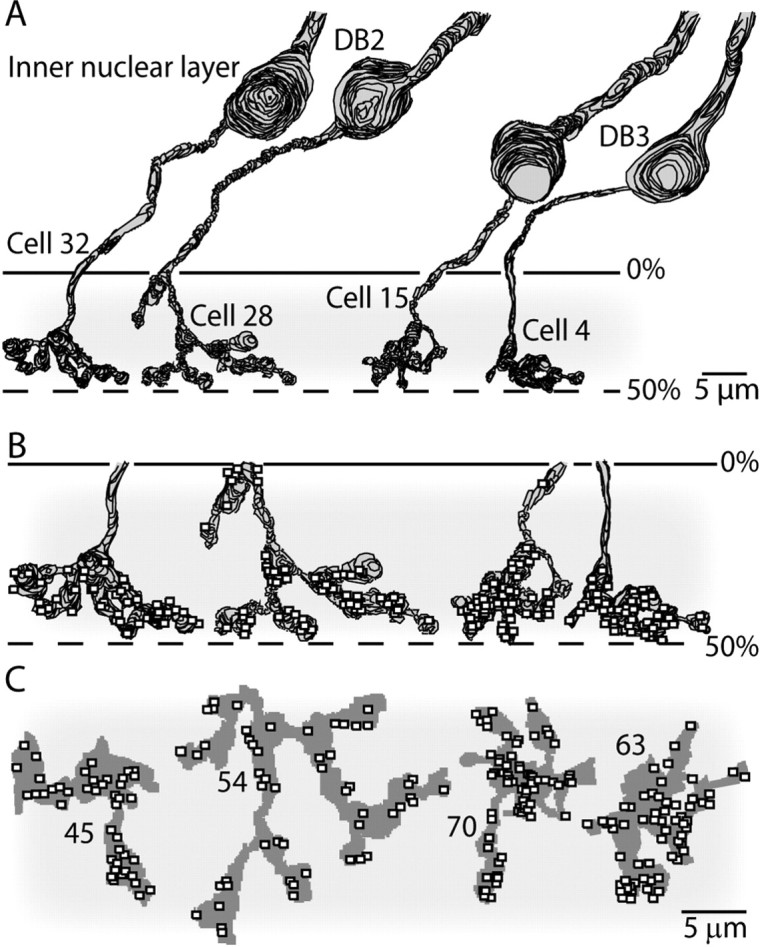
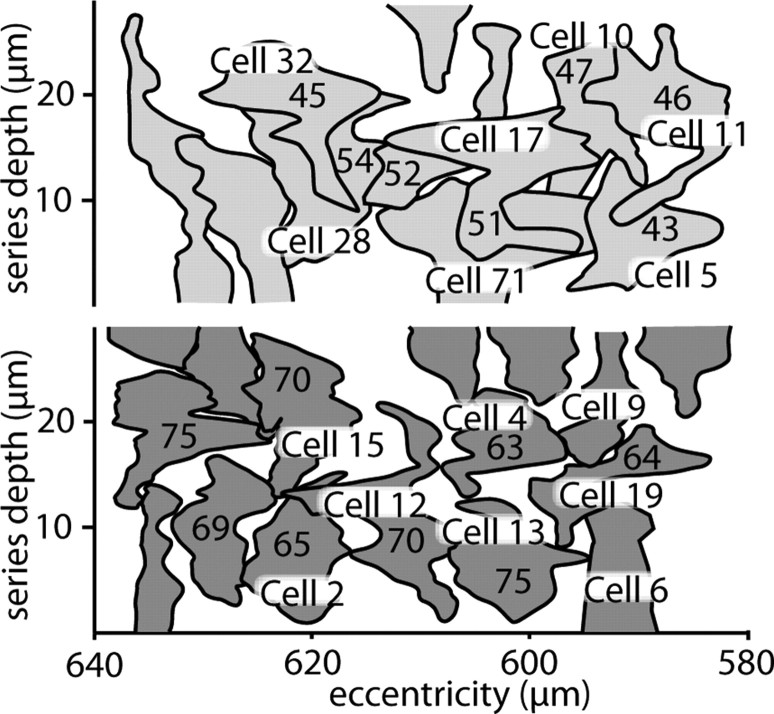
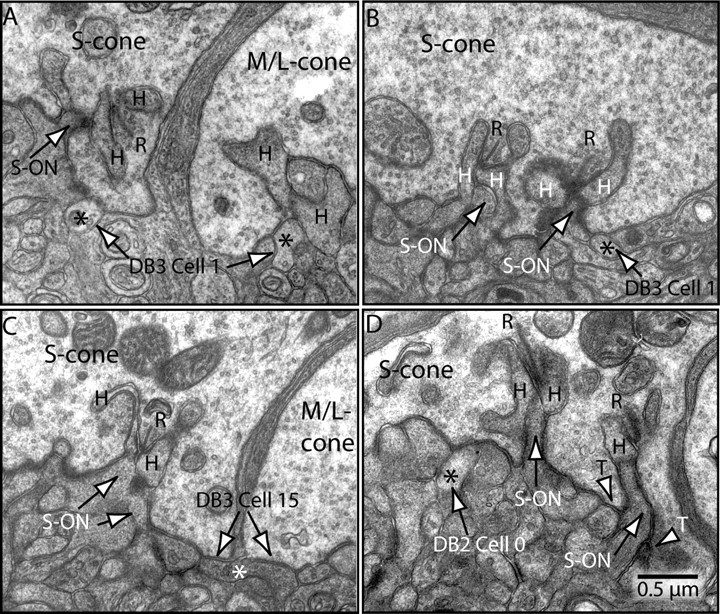
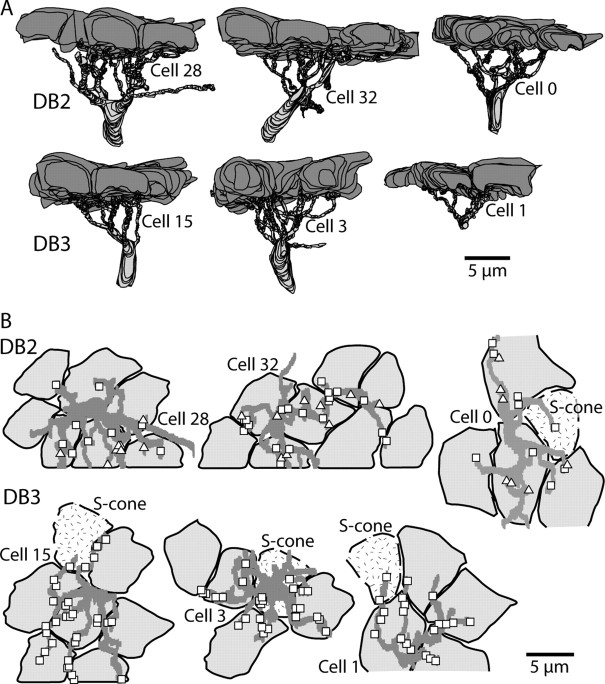
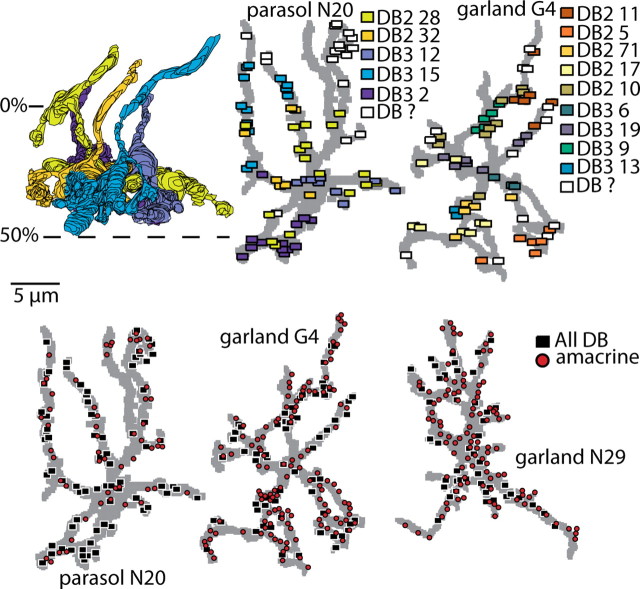
Synaptic circuits in primate fovea have been quantified for midget/parvocellular ganglion cells. Here, based on partial reconstructions from serial electron micrographs, we quantify synaptic circuits for two other types of ganglion cell: the familiar parasol/magnocellular cell and a smaller type, termed "garland." The excitatory circuits both derive from two types of OFF diffuse cone bipolar cell, DB3 and DB2, which collected unselectively from at least 6 +/- 1 cones, including the S type. Cone contacts to DB3 dendrites were usually located between neighboring triads, whereas half of the cone contacts to DB2 were triad associated. Ribbon outputs were as follows: DB3, 69 +/- 5; DB2, 48 +/- 4. A complete parasol cell (30 microm dendritic field diameter) would collect from approximately 50 cones via approximately 120 bipolar and approximately 85 amacrine contacts; a complete garland cell (25 microm dendritic field) would collect from approximately 40 cones via approximately 75 bipolar and approximately 145 amacrine contacts. The bipolar types contributed differently: the parasol cell received most contacts (60%) from DB3, whereas the garland cell received most contacts (67%) from DB2. We hypothesize that DB3 is a transient bipolar cell and that DB2 is sustained. This would be consistent with their relative inputs to the brisk-transient (parasol) ganglion cell. The garland cell, with its high proportion of DB2 inputs plus its high proportion of amacrine synapses (70%) and dense mosaic, might correspond to the local-edge cell in nonprimate retinas, which serves finer acuity at low temporal frequencies. The convergence of S cones onto both types could contribute S-cone input for cortical areas primary visual cortex and the middle temporal area.
Figures
References
-
- Ahmad KM, Klug K, Herr S, Sterling P, Schein S. Cell density ratios in a foveal patch in macaque retina. Vis Neurosci. 2003;20:189–209. - PubMed
-
- Boycott BB, Dowling JE. Organization of the primate retina: light microscopy. Philos Trans R Soc Lond B Biol Sci. 1969;255:109–184. - PubMed
-
- Boycott BB, Wässle H. Morphological classification of bipolar cells of the primate retina. Eur J Neurosci. 1991;3:1069–1088. - PubMed
-
- Calkins DJ. Synaptic organization of cone pathways in the primate retina. In: Gegenfurtner K, Sharpe L, editors. Color vision from molecular genetics to perception. Cambridge, MA: Cambridge UP; 1999.
-
- Calkins DJ. The representation of cone signals in the primate retina. J Opt Soc Am B. 2000;17:597–606. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources