

Sox9 is required for precursor cell expansion and extracellular matrix organization during mouse heart valve development
- PMID: 17350610
- PMCID: PMC1920559
- DOI: 10.1016/j.ydbio.2007.02.002
Sox9 is required for precursor cell expansion and extracellular matrix organization during mouse heart valve development
Abstract
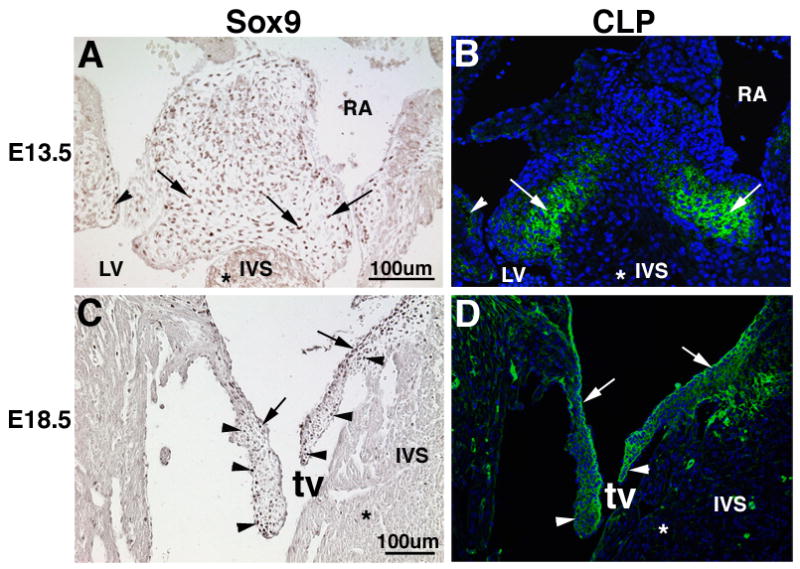
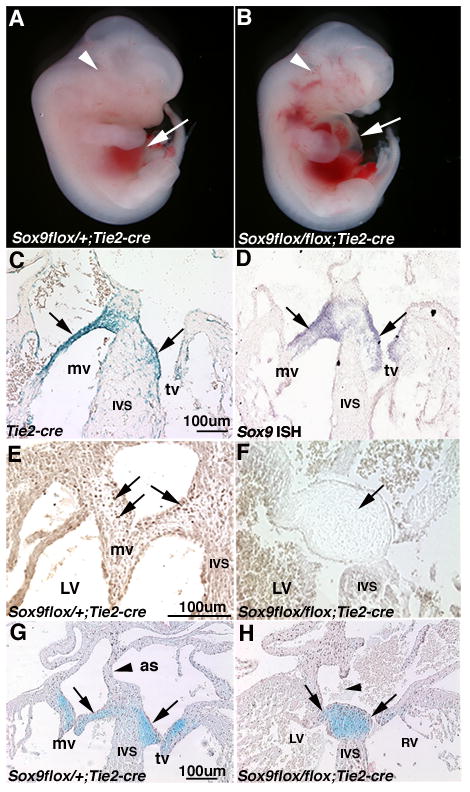
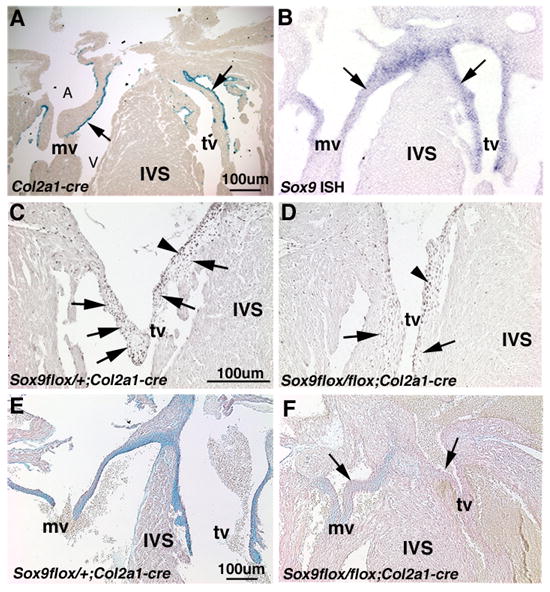
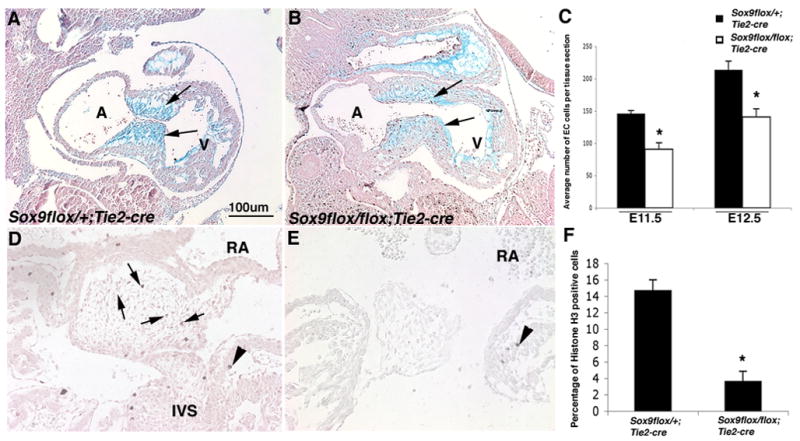
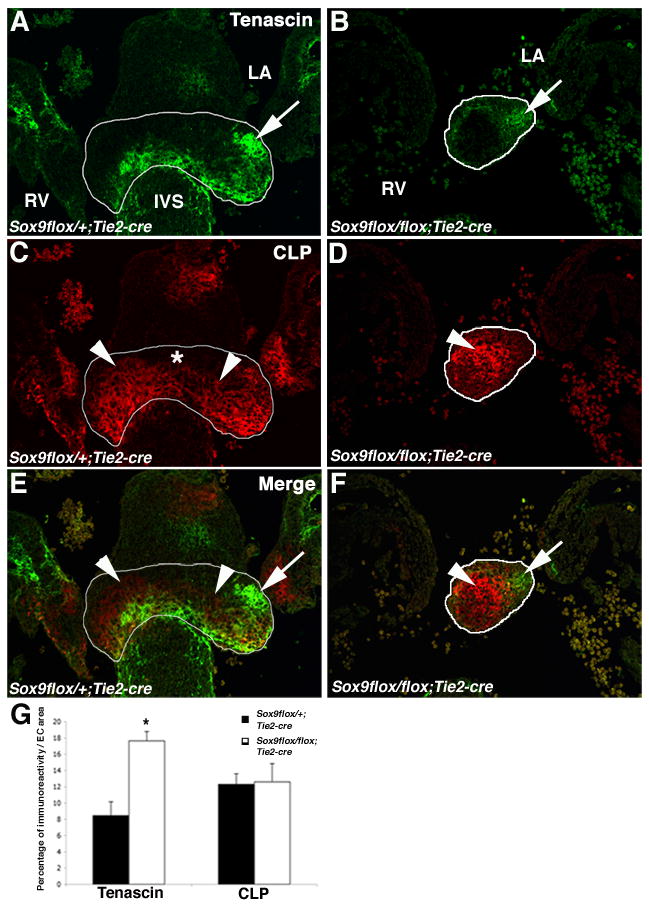
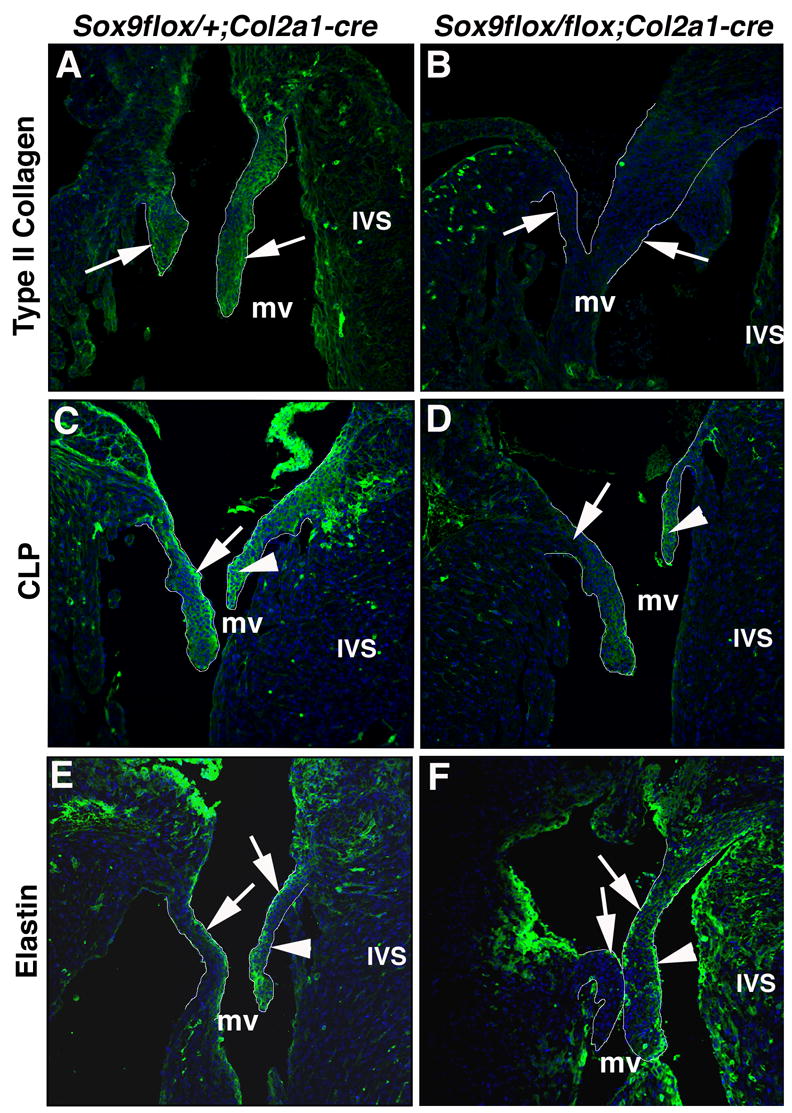
Heart valve structures derived from mesenchymal cells of the endocardial cushions (ECs) are composed of highly organized cell lineages and extracellular matrix. Sox9 is a transcription factor required for both early and late stages of cartilage formation that is also expressed in the developing valves of the heart. The requirements for Sox9 function during valvulogenesis and adult valve homeostasis in mice were examined by conditional inactivation of Sox9 using Tie2-cre and Col2a1-cre transgenes. Sox9(flox/flox);Tie2-cre mice die before E14.5 with hypoplastic ECs, reduced cell proliferation and altered extracellular matrix protein (ECM) deposition. Sox9(flox/flox);Col2a1-cre mice die at birth with thickened heart valve leaflets, reduced expression of cartilage-associated proteins and abnormal ECM patterning. Thickened valve leaflets and calcium deposits, characteristic of valve disease, are observed in heterozygous adult Sox9(flox/+);Col2a1-cre mice. Therefore, Sox9 is required early in valve development for expansion of the precursor cell population and later is required for normal expression and distribution of valvular ECM proteins. These data indicate that Sox9 is required for early and late stages of valvulogenesis and identify a potential role for Sox9 in valve disease mechanisms.
Figures

Similar articles
-
Essential role of Sox9 in the pathway that controls formation of cardiac valves and septa.Proc Natl Acad Sci U S A. 2004 Apr 27;101(17):6502-7. doi: 10.1073/pnas.0401711101. Epub 2004 Apr 19. Proc Natl Acad Sci U S A. 2004. PMID: 15096597 Free PMC article.
-
SOX9 modulates the expression of key transcription factors required for heart valve development.Development. 2015 Dec 15;142(24):4340-50. doi: 10.1242/dev.125252. Epub 2015 Nov 2. Development. 2015. PMID: 26525672
-
Hearts and bones: shared regulatory mechanisms in heart valve, cartilage, tendon, and bone development.Dev Biol. 2006 Jun 15;294(2):292-302. doi: 10.1016/j.ydbio.2006.03.027. Epub 2006 Apr 27. Dev Biol. 2006. PMID: 16643886 Review.
-
Osteo-chondroprogenitor cells are derived from Sox9 expressing precursors.Proc Natl Acad Sci U S A. 2005 Oct 11;102(41):14665-70. doi: 10.1073/pnas.0504750102. Epub 2005 Oct 3. Proc Natl Acad Sci U S A. 2005. PMID: 16203988 Free PMC article.
-
Transcriptional regulation of heart valve progenitor cells.Pediatr Cardiol. 2010 Apr;31(3):414-21. doi: 10.1007/s00246-009-9616-x. Epub 2009 Dec 29. Pediatr Cardiol. 2010. PMID: 20039031 Free PMC article. Review.
Cited by
-
Mechanisms of heart valve development and disease.Development. 2020 Jul 3;147(13):dev183020. doi: 10.1242/dev.183020. Development. 2020. PMID: 32620577 Free PMC article. Review.
-
Twist1 transcriptional targets in the developing atrio-ventricular canal of the mouse.PLoS One. 2012;7(7):e40815. doi: 10.1371/journal.pone.0040815. Epub 2012 Jul 16. PLoS One. 2012. PMID: 22815831 Free PMC article.
-
The Genetic Regulation of Aortic Valve Development and Calcific Disease.Front Cardiovasc Med. 2018 Nov 6;5:162. doi: 10.3389/fcvm.2018.00162. eCollection 2018. Front Cardiovasc Med. 2018. PMID: 30460247 Free PMC article. Review.
-
Sox9 in mouse urogenital sinus epithelium mediates elongation of prostatic buds and expression of genes involved in epithelial cell migration.Gene Expr Patterns. 2019 Dec;34:119075. doi: 10.1016/j.gep.2019.119075. Epub 2019 Oct 25. Gene Expr Patterns. 2019. PMID: 31669249 Free PMC article.
-
Activation of AcvR1-Mediated Signaling Results in Semilunar Valve Defects.J Cardiovasc Dev Dis. 2022 Aug 16;9(8):272. doi: 10.3390/jcdd9080272. J Cardiovasc Dev Dis. 2022. PMID: 36005436 Free PMC article.
References
-
- Baudino T, Carver W, Giles WR, Borg TK. Cardiac Fibroblasts: friend or foe? Am J Physiol Heart Circ Physiol. 2006;291:H1015–26. - PubMed
-
- Bell DM, Leung KK, Wheatley SC, Ng LJ, Zhou S, Ling KW, Sham MH, Koopman P, Tam PP, Cheah KS. SOX9 directly regulates the type-II collagen gene. Nat Genet. 1997;16:174–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
