

Ca2+ stores regulate ryanodine receptor Ca2+ release channels via luminal and cytosolic Ca2+ sites
- PMID: 17351009
- PMCID: PMC1853142
- DOI: 10.1529/biophysj.106.099028
Ca2+ stores regulate ryanodine receptor Ca2+ release channels via luminal and cytosolic Ca2+ sites
Abstract
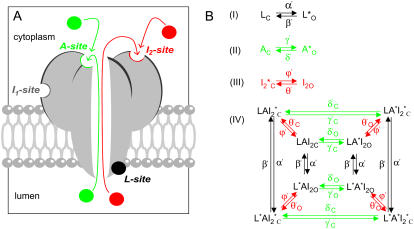
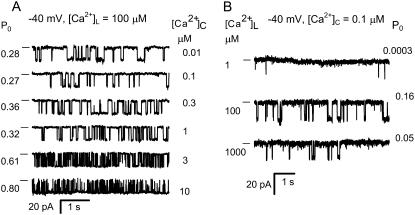
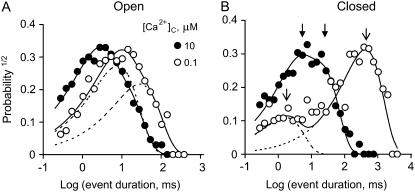
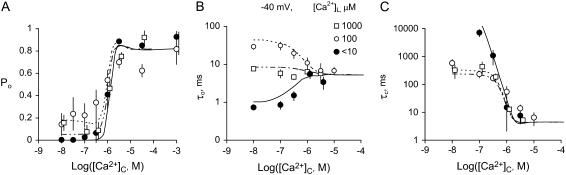
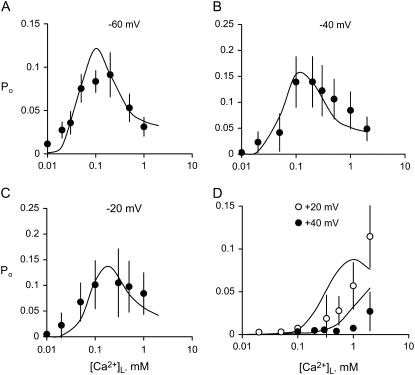
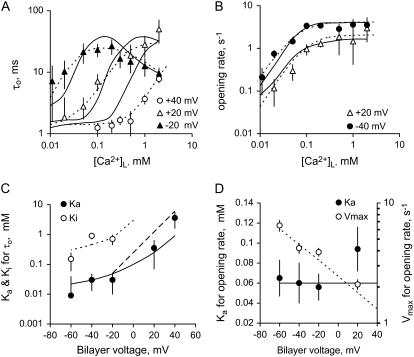
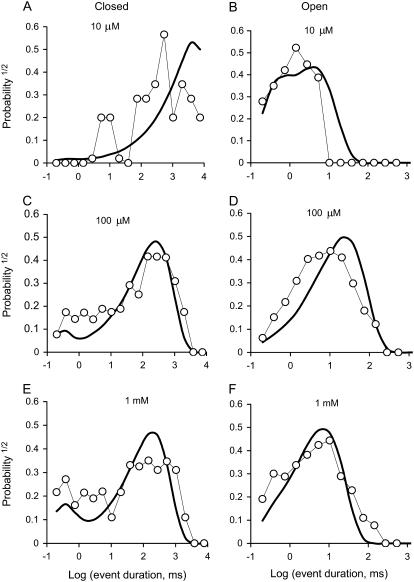
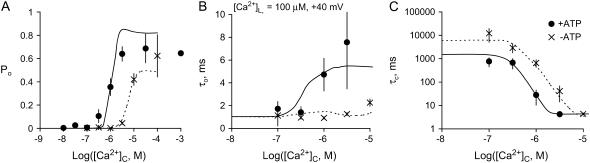
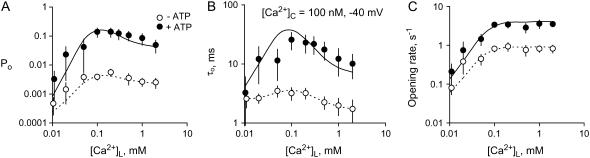
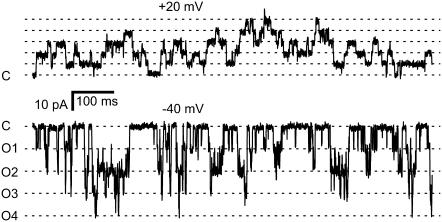
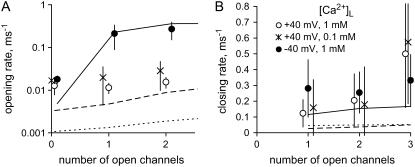
The free [Ca2+] in endoplasmic/sarcoplasmic reticulum Ca2+ stores regulates excitability of Ca2+ release by stimulating the Ca2+ release channels. Just how the stored Ca2+ regulates activation of these channels is still disputed. One proposal attributes luminal Ca2+-activation to luminal facing regulatory sites, whereas another envisages Ca2+ permeation to cytoplasmic sites. This study develops a unified model for luminal Ca2+ activation for single cardiac ryanodine receptors (RyR2) and RyRs in coupled clusters in artificial lipid bilayers. It is shown that luminal regulation of RyR2 involves three modes of action associated with Ca2+ sensors in different parts of the molecule; a luminal activation site (L-site, 60 microM affinity), a cytoplasmic activation site (A-site, 0.9 microM affinity), and a novel cytoplasmic inactivation site (I2-site, 1.2 microM affinity). RyR activation by luminal Ca2+ is demonstrated to occur by a multistep process dubbed luminal-triggered Ca2+ feedthrough. Ca2+ binding to the L-site initiates brief openings (1 ms duration at 1-10 s(-1)) allowing luminal Ca2+ to access the A-site, producing up to 30-fold prolongation of openings. The model explains a broad data set, reconciles previous conflicting observations and provides a foundation for understanding the action of pharmacological agents, RyR-associated proteins, and RyR2 mutations on a range of Ca2+-mediated physiological and pathological processes.
Figures
References
-
- Stern, M. D., and H. Cheng. 2004. Putting out the fire: what terminates calcium-induced calcium release in cardiac muscle? Cell Calcium. 35:591–601. - PubMed
-
- Coombes, S., R. Hinch, and Y. Timofeeva. 2004. Receptors, sparks and waves in a fire-diffuse-fire framework for calcium release. Prog. Biophys. Mol. Biol. 85:197–216. - PubMed
-
- Fabiato, A., and F. Fabiato. 1977. Calcium release from the sarcoplasmic reticulum. Circ. Res. 40:119–129. - PubMed
-
- Verkhratsky, A. 2005. Physiology and pathophysiology of the calcium store in the endoplasmic reticulum of neurons. Physiol. Rev. 85:201–279. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
