

VanA-type enterococci from humans, animals, and food: species distribution, population structure, Tn1546 typing and location, and virulence determinants
- PMID: 17351100
- PMCID: PMC1907105
- DOI: 10.1128/AEM.02239-06
VanA-type enterococci from humans, animals, and food: species distribution, population structure, Tn1546 typing and location, and virulence determinants
Abstract
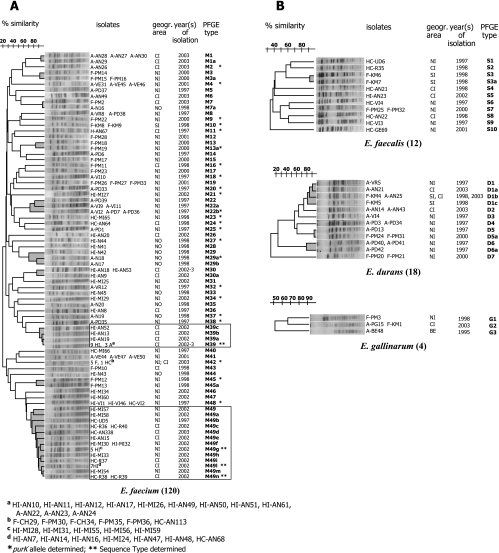
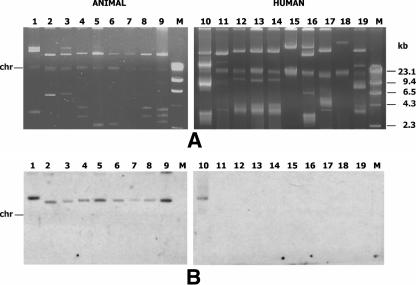
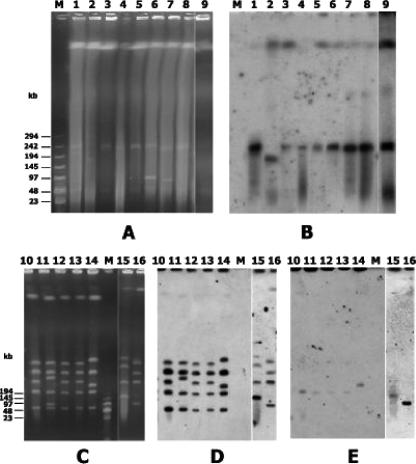
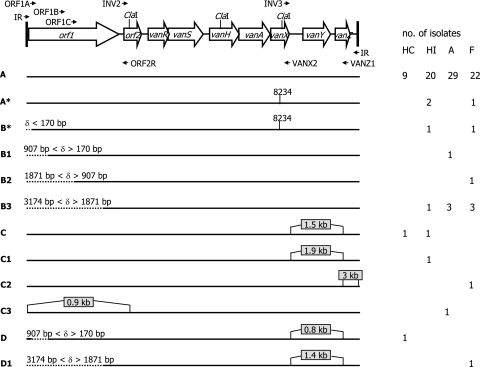
VanA-type human (n=69), animal (n=49), and food (n=36) glycopeptide-resistant enterococci (GRE) from different geographic areas were investigated to study their possible reservoirs and transmission routes. Pulsed-field gel electrophoresis (PFGE) revealed two small genetically related clusters, M39 (n=4) and M49 (n=13), representing Enterococcus faecium isolates from animal and human feces and from clinical and fecal human samples. Multilocus sequence typing showed that both belonged to the epidemic lineage of CC17. purK allele analysis of 28 selected isolates revealed that type 1 was prevalent in human strains (8/11) and types 6 and 3 (14/15) were prevalent in poultry (animals and meat). One hundred and five of the 154 VanA GRE isolates, encompassing different species, origins, and PFGE types, were examined for Tn1546 type and location (plasmid or chromosome) and the incidence of virulence determinants. Hybridization of S1- and I-CeuI-digested total DNA revealed a plasmid location in 98% of the isolates. Human intestinal and animal E. faecium isolates bore large (>150 kb) vanA plasmids. Results of PCR-restriction fragment length polymorphism and sequencing showed the presence of prototype Tn1546 in 80% of strains and the G-to-T mutation at position 8234 in three human intestinal and two pork E. faecium isolates. There were no significant associations (P>0.5) between Tn1546 type and GRE source or enterococcal species. Virulence determinants were detected in all reservoirs but were significantly more frequent (P<0.02) among clinical strains. Multiple determinants were found in clinical and meat Enterococcus faecalis isolates. The presence of indistinguishable vanA elements (mostly plasmid borne) and virulence determinants in different species and PFGE-diverse populations in the presence of host-specific purK housekeeping genes suggested that all GRE might be potential reservoirs of resistance determinants and virulence traits transferable to human-adapted clusters.
Figures
References
-
- Arthur, M., P. E. Reynolds, F. Depardieu, S. Evers, S. Dutka Malen, R. J. Quintiliani, and P. Courvalin. 1996. Mechanisms of glycopeptide resistance in enterococci. J. Infect. 32:11-16. - PubMed
-
- Biavasco, F., A. Miele, C. Vignaroli, E. Manso, R. Lupidi, and P. E. Varaldo. 1996. Genotypic characterization of a nosocomial outbreak of vanA Enterococcus faecalis. Microb. Drug Res. 2:231-237. - PubMed
-
- Biavasco, F., C. Paladini, C. Vignaroli, G. Foglia, E. Manso, and P. E. Varaldo. 2001. Recovery from a single blood culture of two Enterococcus gallinarum isolates carrying both vanC-1 and vanA cluster genes and differing in glycopeptide susceptibility. Eur. J. Clin. Microbiol. Infect. Dis. 20:309-314. - PubMed
-
- Bonora, M. G., C. Bordin, L. Bragagnolo, L. Girelli, M. De Fatima, A. Grossato, M. Ligozzi, G. Lo Cascio, and R. Fontana. 2001. Molecular analysis of vanA enterococci isolated from humans and animals in northeastern Italy. Microb. Drug Resist. 7:247-255. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
