

Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance
- PMID: 17351116
- PMCID: PMC1867366
- DOI: 10.1105/tpc.106.048058
Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance
Abstract
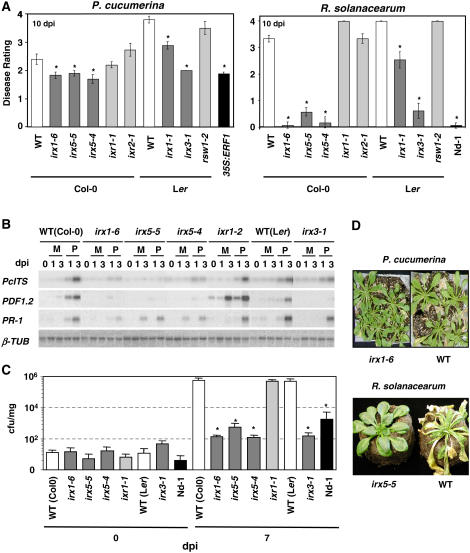
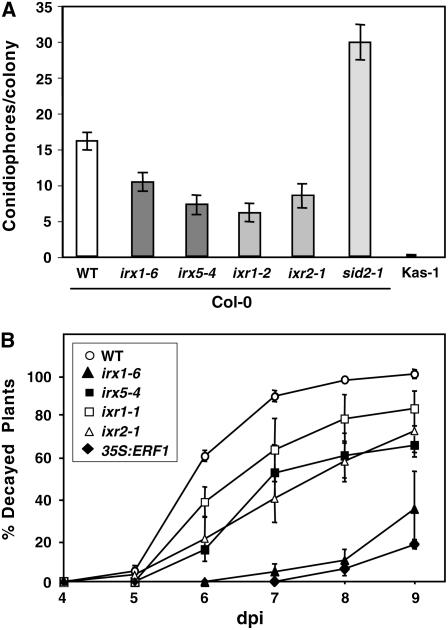
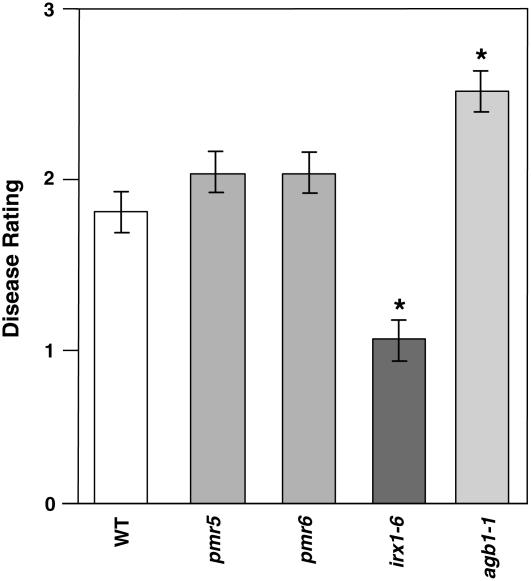
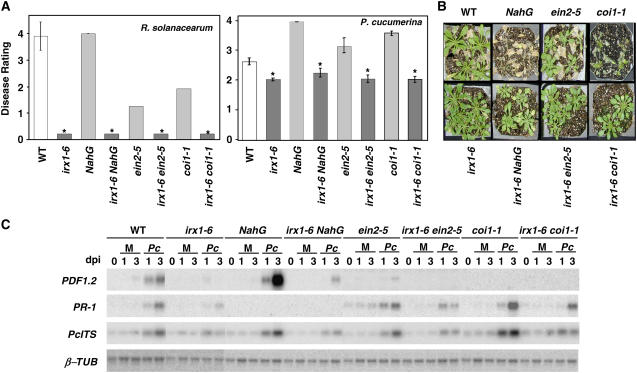
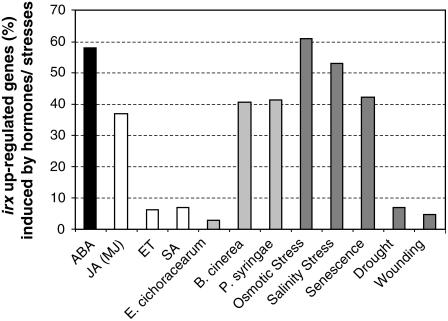
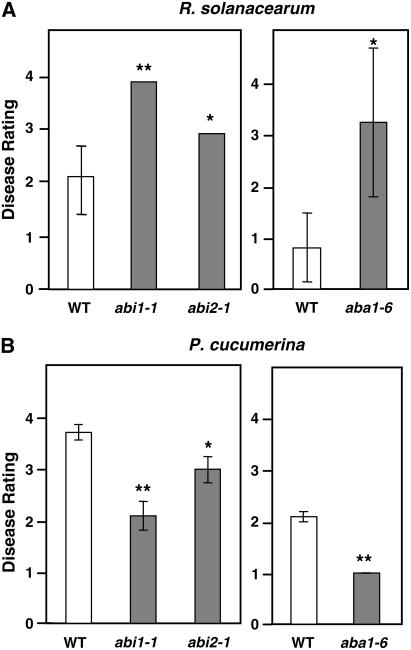
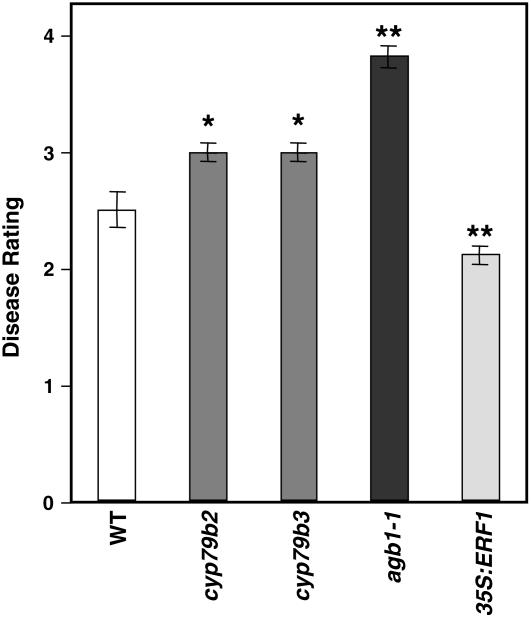
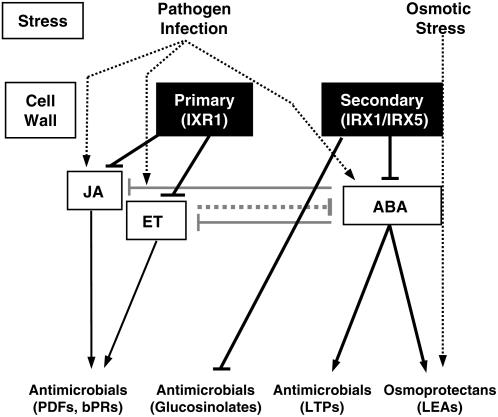
Cellulose is synthesized by cellulose synthases (CESAs) contained in plasma membrane-localized complexes. In Arabidopsis thaliana, three types of CESA subunits (CESA4/IRREGULAR XYLEM5 [IRX5], CESA7/IRX3, and CESA8/IRX1) are required for secondary cell wall formation. We report that mutations in these proteins conferred enhanced resistance to the soil-borne bacterium Ralstonia solanacearum and the necrotrophic fungus Plectosphaerella cucumerina. By contrast, susceptibility to these pathogens was not altered in cell wall mutants of primary wall CESA subunits (CESA1, CESA3/ISOXABEN RESISTANT1 [IXR1], and CESA6/IXR2) or POWDERY MILDEW-RESISTANT5 (PMR5) and PMR6 genes. Double mutants indicated that irx-mediated resistance was independent of salicylic acid, ethylene, and jasmonate signaling. Comparative transcriptomic analyses identified a set of common irx upregulated genes, including a number of abscisic acid (ABA)-responsive, defense-related genes encoding antibiotic peptides and enzymes involved in the synthesis and activation of antimicrobial secondary metabolites. These data as well as the increased susceptibility of ABA mutants (abi1-1, abi2-1, and aba1-6) to R. solanacearum support a direct role of ABA in resistance to this pathogen. Our results also indicate that alteration of secondary cell wall integrity by inhibiting cellulose synthesis leads to specific activation of novel defense pathways that contribute to the generation of an antimicrobial-enriched environment hostile to pathogens.
Figures
References
-
- AbuQamar, S., Chen, X., Dhawan, R., Bluhm, B., Salmeron, J., Lam, S., Dietrich, R.A., and Mengiste, T. (2006). Expression profiling and mutant analysis reveals complex regulatory networks involved in Arabidopsis response to Botrytis infection. Plant J. 48 28–44. - PubMed
-
- Anderson, J.P., Badruzsaufari, E., Schenk, P.M., Manners, J.M., Desmond, O.J., Ehlert, C., Maclean, D.J., Ebert, P.R., and Kazan, K. (2004). Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 16 3460–3479. - PMC - PubMed
-
- Bartsch, M., Gobbato, E., Bednarek, P., Debey, S., Schultze, J.L., Bautor, J., and Parker, J.E. (2006). Salicylic acid-independent ENHANCED DISEASE SUSCEPTIBILITY1 signaling in Arabidopsis immunity and cell death is regulated by the monooxygenase FMO1 and the Nudix hydrolase NUDT7. Plant Cell 18 1038–1051. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
