

Repetitive DNA is associated with centromeric domains in Trypanosoma brucei but not Trypanosoma cruzi
- PMID: 17352808
- PMCID: PMC1868937
- DOI: 10.1186/gb-2007-8-3-r37
Repetitive DNA is associated with centromeric domains in Trypanosoma brucei but not Trypanosoma cruzi
Abstract
Background: Trypanosomes are parasitic protozoa that diverged early from the main eukaryotic lineage. Their genomes display several unusual characteristics and, despite completion of the trypanosome genome projects, the location of centromeric DNA has not been identified.
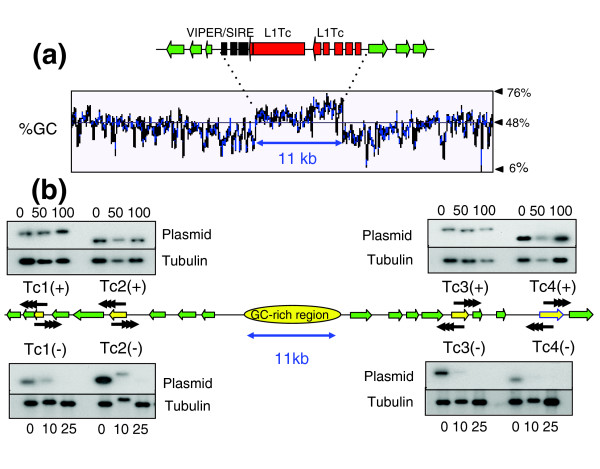
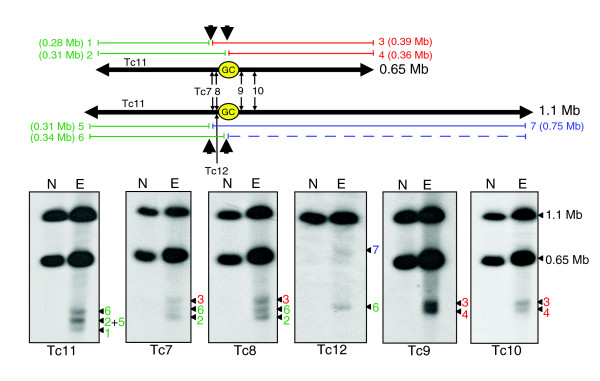
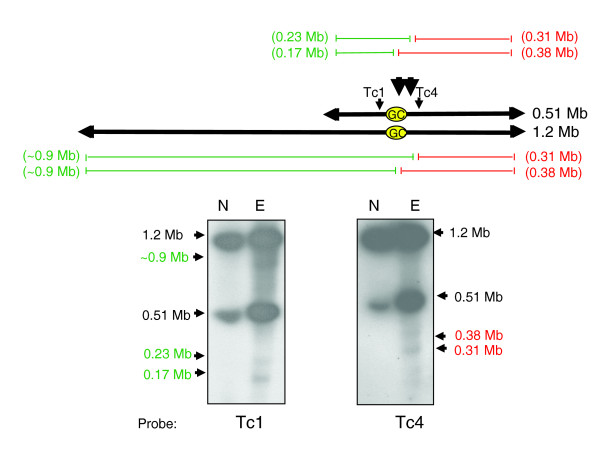
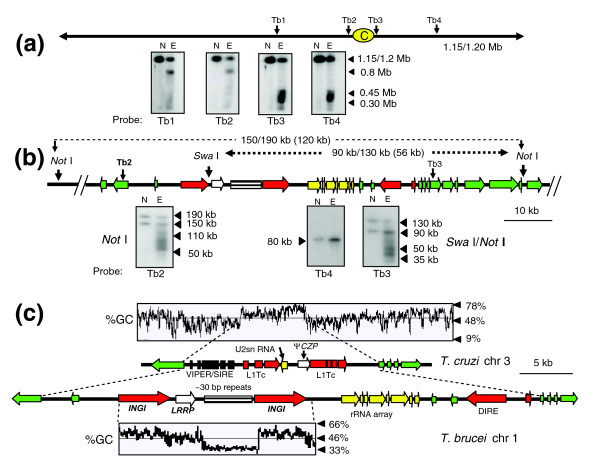
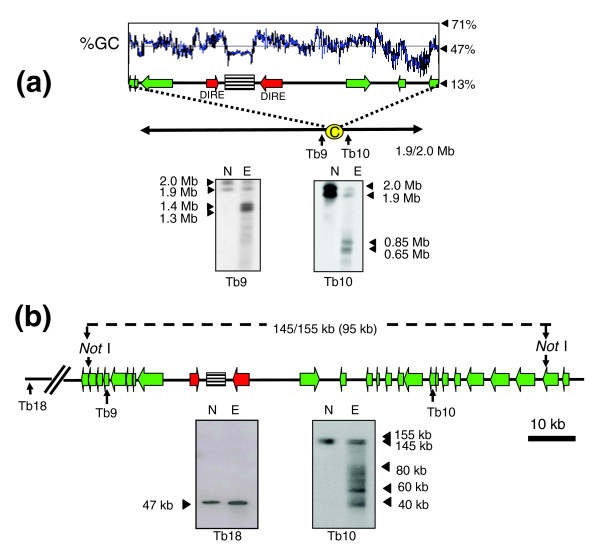
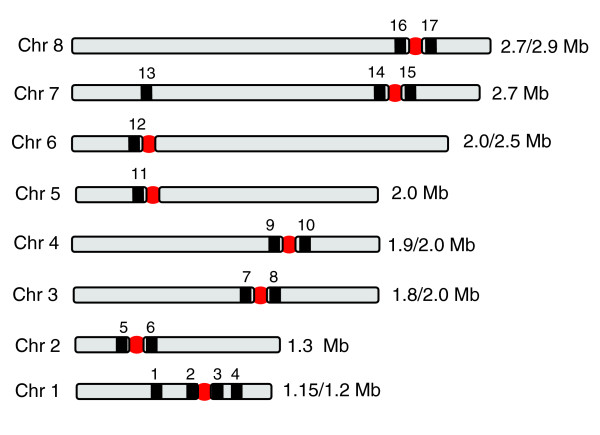
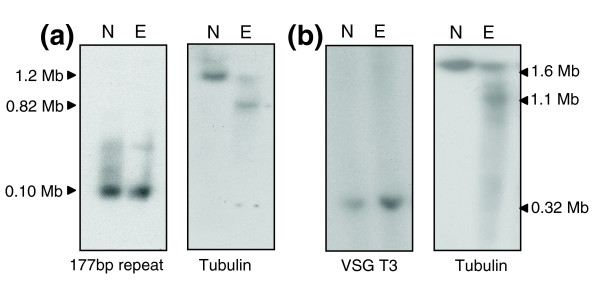
Results: We report evidence on the location and nature of centromeric DNA in Trypanosoma cruzi and Trypanosoma brucei. In T. cruzi, we used telomere-associated chromosome fragmentation and found that GC-rich transcriptional 'strand-switch' domains composed predominantly of degenerate retrotranposons are a shared feature of regions that confer mitotic stability. Consistent with this, etoposide-mediated topoisomerase-II cleavage, a biochemical marker for active centromeres, is concentrated at these domains. In the 'megabase-sized' chromosomes of T. brucei, topoisomerase-II activity is also focused at single loci that encompass regions between directional gene clusters that contain transposable elements. Unlike T. cruzi, however, these loci also contain arrays of AT-rich repeats stretching over several kilobases. The sites of topoisomerase-II activity on T. brucei chromosome 1 and T. cruzi chromosome 3 are syntenic, suggesting that centromere location has been conserved for more than 200 million years. The T. brucei intermediate and minichromosomes, which lack housekeeping genes, do not exhibit site-specific accumulation of topoisomerase-II, suggesting that segregation of these atypical chromosomes might involve a centromere-independent mechanism.
Conclusion: The localization of centromeric DNA in trypanosomes fills a major gap in our understanding of genome organization in these important human pathogens. These data are a significant step towards identifying and functionally characterizing other determinants of centromere function and provide a framework for dissecting the mechanisms of chromosome segregation.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
