

Requirement of ZO-1 for the formation of belt-like adherens junctions during epithelial cell polarization
- PMID: 17353356
- PMCID: PMC2064052
- DOI: 10.1083/jcb.200612080
Requirement of ZO-1 for the formation of belt-like adherens junctions during epithelial cell polarization
Abstract
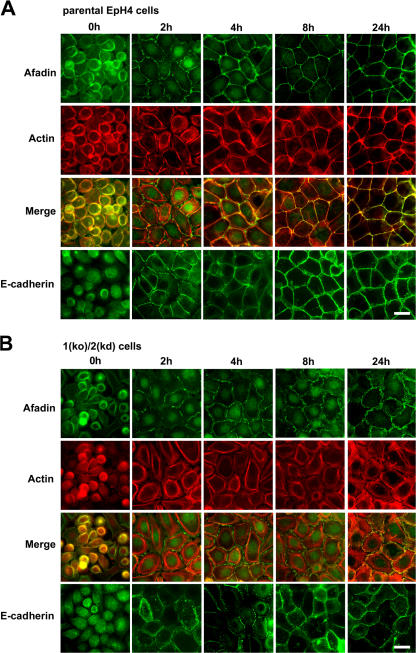
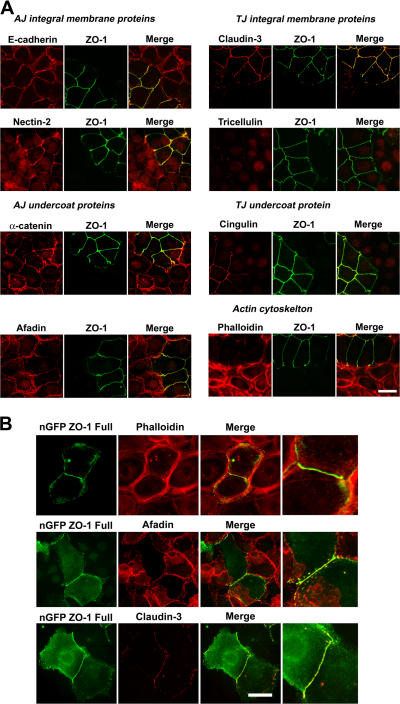
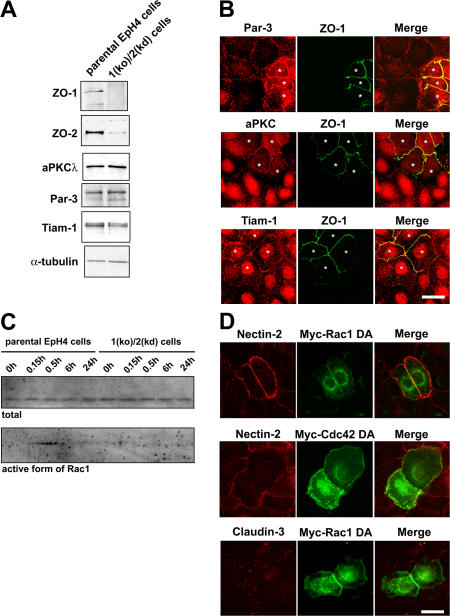
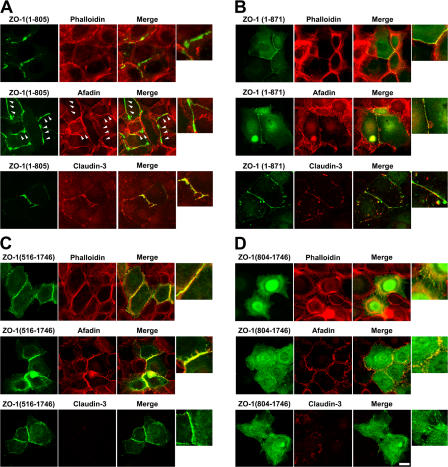
The molecular mechanisms of how primordial adherens junctions (AJs) evolve into spatially separated belt-like AJs and tight junctions (TJs) during epithelial polarization are not well understood. Previously, we reported the establishment of ZO-1/ZO-2-deficient cultured epithelial cells (1[ko]/2[kd] cells), which lacked TJs completely. In the present study, we found that the formation of belt-like AJs was significantly delayed in 1(ko)/2(kd) cells during epithelial polarization. The activation of Rac1 upon primordial AJ formation is severely impaired in 1(ko)/2(kd) cells. Our data indicate that ZO-1 plays crucial roles not only in TJ formation, but also in the conversion from "fibroblastic" AJs to belt-like "polarized epithelial" AJs through Rac1 activation. Furthermore, to examine whether ZO-1 itself mediate belt-like AJ and TJ formation, respectively, we performed a mutational analysis of ZO-1. The requirement for ZO-1 differs between belt-like AJ and TJ formation. We propose that ZO-1 is directly involved in the establishment of two distinct junctional domains, belt-like AJs and TJs, during epithelial polarization.
Figures

References
-
- Anderson, J.M., C.M. Van Itallie, and A.S. Fanning. 2004. Setting up a selective barrier at the apical junction complex. Curr. Opin. Cell Biol. 16:140–145. - PubMed
-
- Ando-Akatsuka, Y., S. Yonemura, M. Itoh, M. Furuse, and S. Tsukita. 1999. Differential behavior of E-cadherin and occludin in their colocalization with ZO-1 during the establishment of epithelial cell polarity. J. Cell. Physiol. 179:115–125. - PubMed
-
- Chen, X., and I.G. Macara. 2005. Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat. Cell Biol. 7:262–269. - PubMed
-
- Furuse, M., and S. Tsukita. 2006. Claudins in occluding junctions of humans and flies. Trends Cell Biol. 16:181–188. - PubMed
-
- Gonzalez-Mariscal, L., A. Betanzos, and A. Avila-Flores. 2000. MAGUK proteins: structure and role in the tight junction. Semin. Cell Dev. Biol. 11:315–324. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
