

IL-31-IL-31R interactions negatively regulate type 2 inflammation in the lung
- PMID: 17353366
- PMCID: PMC2137900
- DOI: 10.1084/jem.20061791
IL-31-IL-31R interactions negatively regulate type 2 inflammation in the lung
Abstract
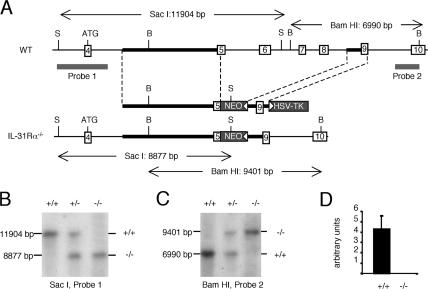
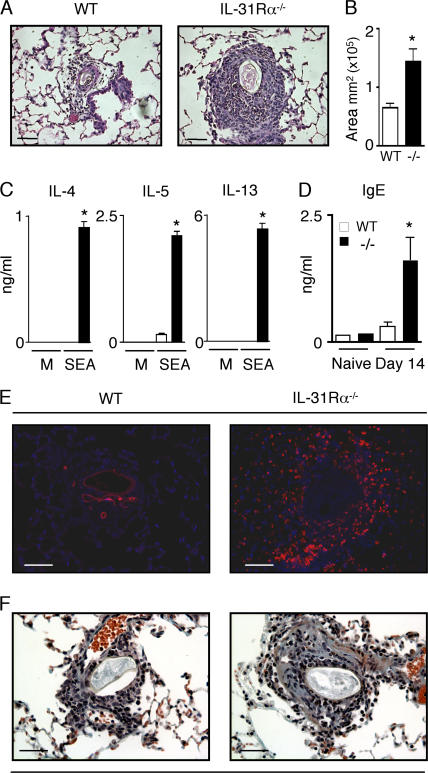
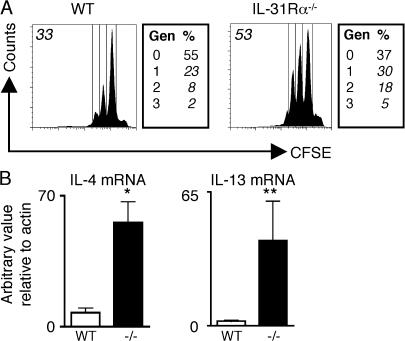
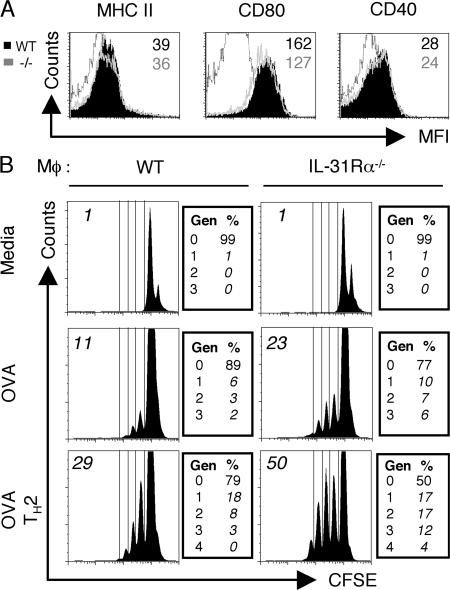
Interleukin (IL) 31Ralpha (glycoprotein 130-like monocyte receptor and glycoprotein 130-like receptor) heterodimerizes with oncostatin M receptor beta to bind IL-31, a cytokine expressed preferentially by CD4(+) T helper type 2 (Th2) cells. However, the functions of IL-31-IL-31R signaling in immune regulation remain unknown. Here, we identify a novel role for IL-31R in limiting type 2 inflammation in the lung. After intravenous injection of Schistosoma mansoni eggs, IL-31Ralpha(-/-) mice developed severe pulmonary inflammation, characterized by an increase in the area of granulomatous inflammation, increased numbers of resistin-like molecule alpha(+) cells, and enhanced collagen deposition compared to WT counterparts. In vitro, macrophages generated from IL-31Ralpha(-/-) mice promoted enhanced ovalbumin-specific CD4(+) T cell proliferation and purified naive CD4(+) T cells from IL-31Ralpha(-/-) mice exhibited enhanced proliferation and expression of Th2 cytokines, identifying a T cell- and macrophage-intrinsic regulatory function for IL-31R signaling. In contrast, the generation of CD4(+) T cell-mediated Th1 responses were normal in IL-31Ralpha(-/-) mice, suggesting that the regulatory role of IL-31R signaling is limited to type 2 responses. Together, these data implicate IL-31R signaling as a novel negative regulatory pathway that specifically limits type 2 inflammation.
Figures
References
-
- Boulay, J.L., J.J. O'Shea, and W.E. Paul. 2003. Molecular phylogeny within type I cytokines and their cognate receptors. Immunity. 19:159– 163. - PubMed
-
- Hofmann, S.R., R. Ettinger, Y.J. Zhou, M. Gadina, P. Lipsky, R. Siegel, F. Candotti, and J.J. O'Shea. 2002. Cytokines and their role in lymphoid development, differentiation and homeostasis. Curr. Opin. Allergy Clin. Immunol. 2:495–506. - PubMed
-
- Hunter, C.A. 2005. New IL-12-family members: IL-23 and IL-27, cytokines with divergent functions. Nat. Rev. Immunol. 5:521–531. - PubMed
-
- Dreuw, A., S. Radtke, S. Pflanz, B.E. Lippok, P.C. Heinrich, and H.M. Hermanns. 2004. Characterization of the signaling capacities of the novel gp130-like cytokine receptor. J. Biol. Chem. 279:36112–36120. - PubMed
-
- Diveu, C., E. Lelievre, D. Perret, A.H. Lak-Hal, J. Froger, C. Guillet, S. Chevalier, F. Rousseau, A. Wesa, L. Preisser, et al. 2003. GPL, a novel cytokine receptor related to GP130 and leukemia inhibitory factor receptor. J. Biol. Chem. 278:49850–49859. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
