

An aromatic sensor with aversion to damaged strands confers versatility to DNA repair
- PMID: 17355181
- PMCID: PMC1820611
- DOI: 10.1371/journal.pbio.0050079
An aromatic sensor with aversion to damaged strands confers versatility to DNA repair
Abstract
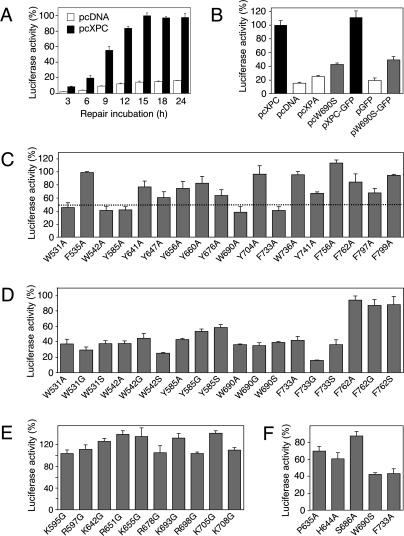
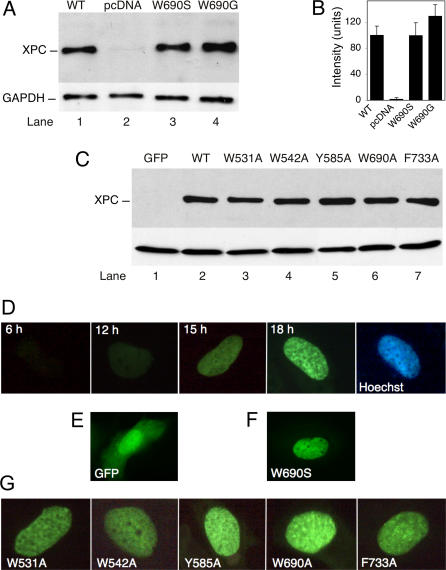
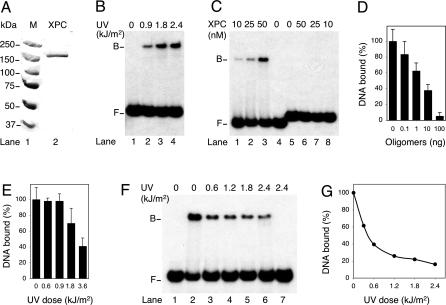
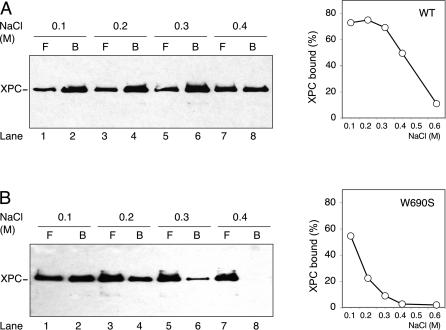
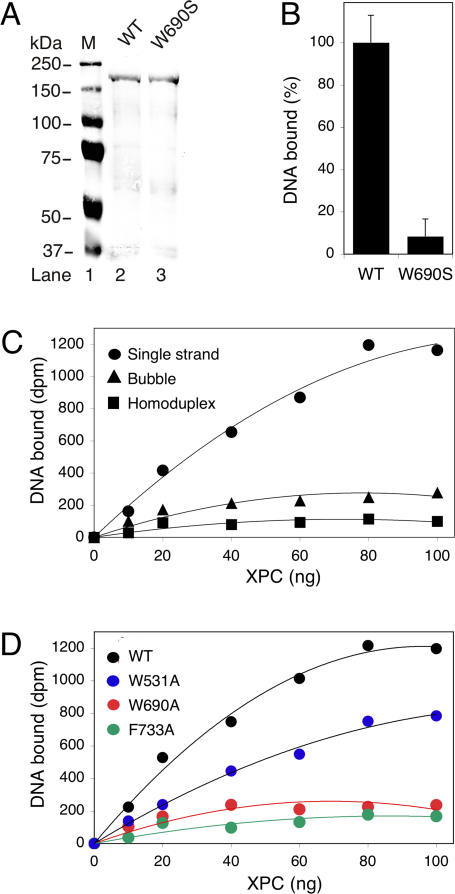
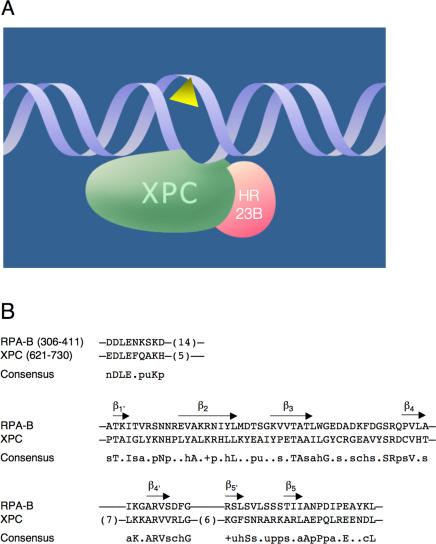
It was not known how xeroderma pigmentosum group C (XPC) protein, the primary initiator of global nucleotide excision repair, achieves its outstanding substrate versatility. Here, we analyzed the molecular pathology of a unique Trp690Ser substitution, which is the only reported missense mutation in xeroderma patients mapping to the evolutionary conserved region of XPC protein. The function of this critical residue and neighboring conserved aromatics was tested by site-directed mutagenesis followed by screening for excision activity and DNA binding. This comparison demonstrated that Trp690 and Phe733 drive the preferential recruitment of XPC protein to repair substrates by mediating an exquisite affinity for single-stranded sites. Such a dual deployment of aromatic side chains is the distinctive feature of functional oligonucleotide/oligosaccharide-binding folds and, indeed, sequence homologies with replication protein A and breast cancer susceptibility 2 protein indicate that XPC displays a monomeric variant of this recurrent interaction motif. An aversion to associate with damaged oligonucleotides implies that XPC protein avoids direct contacts with base adducts. These results reveal for the first time, to our knowledge, an entirely inverted mechanism of substrate recognition that relies on the detection of single-stranded configurations in the undamaged complementary sequence of the double helix.
Conflict of interest statement
Figures


Similar articles
-
In vivo destabilization and functional defects of the xeroderma pigmentosum C protein caused by a pathogenic missense mutation.Mol Cell Biol. 2007 Oct;27(19):6606-14. doi: 10.1128/MCB.02166-06. Epub 2007 Aug 6. Mol Cell Biol. 2007. PMID: 17682058 Free PMC article.
-
Dissection of the xeroderma pigmentosum group C protein function by site-directed mutagenesis.Antioxid Redox Signal. 2011 Jun 15;14(12):2479-90. doi: 10.1089/ars.2010.3399. Epub 2010 Oct 7. Antioxid Redox Signal. 2011. PMID: 20649465
-
Xeroderma pigmentosum complementation group A protein is driven to nucleotide excision repair sites by the electrostatic potential of distorted DNA.DNA Repair (Amst). 2007 Dec 1;6(12):1819-28. doi: 10.1016/j.dnarep.2007.07.011. Epub 2007 Aug 31. DNA Repair (Amst). 2007. PMID: 17765667
-
Dynamic two-stage mechanism of versatile DNA damage recognition by xeroderma pigmentosum group C protein.Mutat Res. 2010 Mar 1;685(1-2):21-8. doi: 10.1016/j.mrfmmm.2009.08.005. Epub 2009 Aug 15. Mutat Res. 2010. PMID: 19686765 Review.
-
Mechanisms of DNA damage recognition and strand discrimination in human nucleotide excision repair.DNA Repair (Amst). 2004 Nov 2;3(11):1409-23. doi: 10.1016/j.dnarep.2004.05.005. DNA Repair (Amst). 2004. PMID: 15380097 Review.
Cited by
-
Xeroderma pigmentosum complementation group C protein (XPC) serves as a general sensor of damaged DNA.DNA Repair (Amst). 2013 Nov;12(11):947-53. doi: 10.1016/j.dnarep.2013.08.013. Epub 2013 Sep 17. DNA Repair (Amst). 2013. PMID: 24051049 Free PMC article.
-
A chromatin scaffold for DNA damage recognition: how histone methyltransferases prime nucleosomes for repair of ultraviolet light-induced lesions.Nucleic Acids Res. 2020 Feb 28;48(4):1652-1668. doi: 10.1093/nar/gkz1229. Nucleic Acids Res. 2020. PMID: 31930303 Free PMC article.
-
Nucleotide excision repair leaves a mark on chromatin: DNA damage detection in nucleosomes.Cell Mol Life Sci. 2021 Dec;78(24):7925-7942. doi: 10.1007/s00018-021-03984-7. Epub 2021 Nov 3. Cell Mol Life Sci. 2021. PMID: 34731255 Free PMC article. Review.
-
Two-stage dynamic DNA quality check by xeroderma pigmentosum group C protein.EMBO J. 2009 Aug 19;28(16):2387-99. doi: 10.1038/emboj.2009.187. Epub 2009 Jul 16. EMBO J. 2009. PMID: 19609301 Free PMC article.
-
Low-intensity red and infrared lasers affect mRNA expression of DNA nucleotide excision repair in skin and muscle tissue.Lasers Med Sci. 2016 Apr;31(3):429-35. doi: 10.1007/s10103-016-1870-6. Epub 2016 Jan 21. Lasers Med Sci. 2016. PMID: 26796702
References
-
- Sancar A. DNA excision repair. Annu Rev Biochem. 1996;65:43–81. - PubMed
-
- Wood RD. Nucleotide excision repair in mammalian cells. J Biol Chem. 1997;272:23465–23468. - PubMed
-
- De Laat WL, Jaspers NG, Hoeijmakers JHJ. Molecular mechanism of nucleotide excision repair. Genes Dev. 1999;13:768–785. - PubMed
-
- Gillet LC, Schärer OD. Molecular mechanisms of mammalian global genome nucleotide excision repair. Chem Rev. 2006;106:253–276. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
