

Coordination of steps in single-nucleotide base excision repair mediated by apurinic/apyrimidinic endonuclease 1 and DNA polymerase beta
- PMID: 17355977
- PMCID: PMC2366199
- DOI: 10.1074/jbc.M611295200
Coordination of steps in single-nucleotide base excision repair mediated by apurinic/apyrimidinic endonuclease 1 and DNA polymerase beta
Abstract
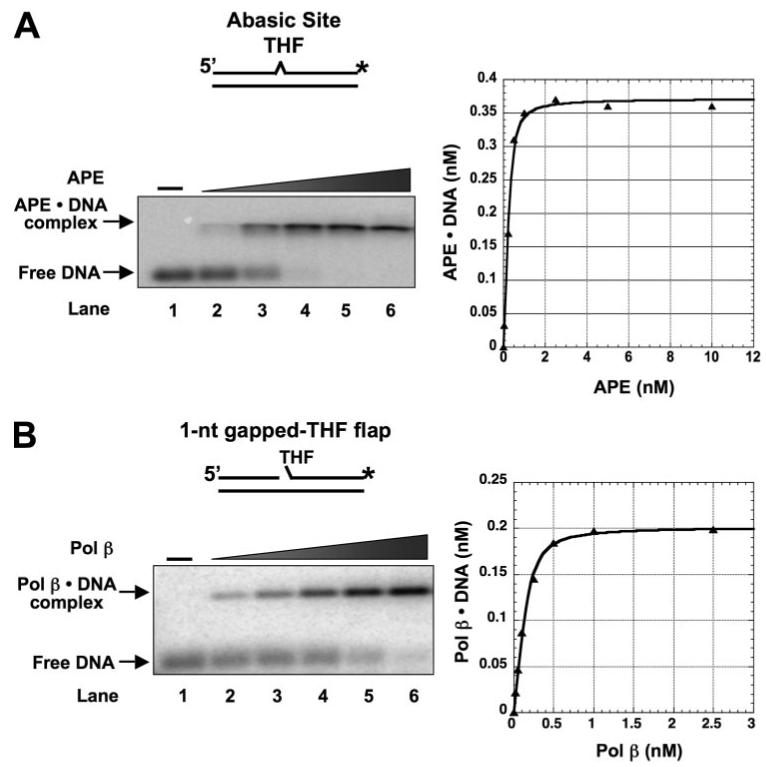
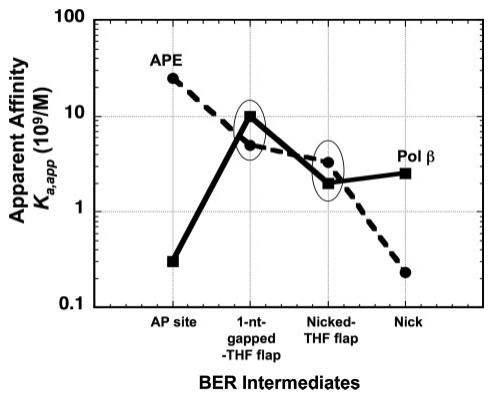
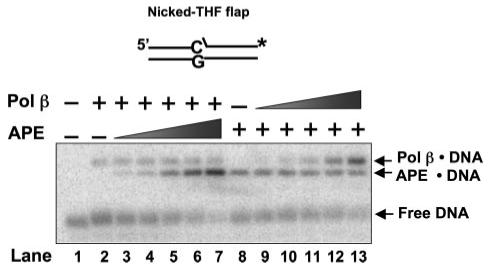
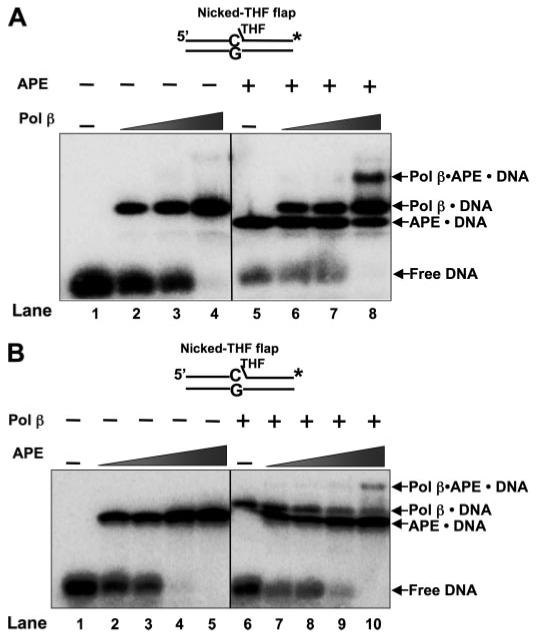
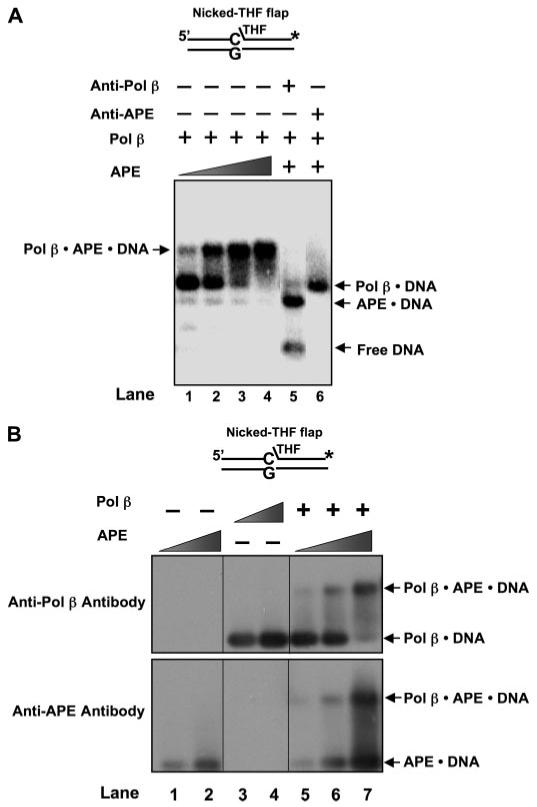
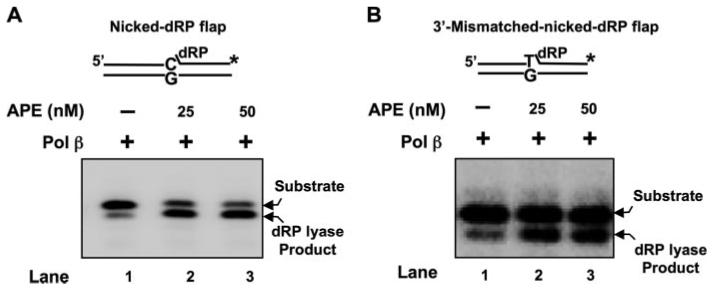
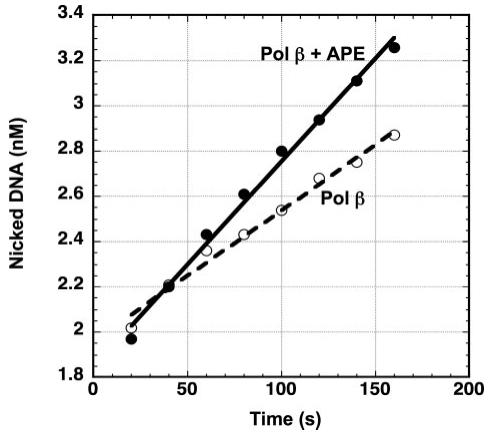
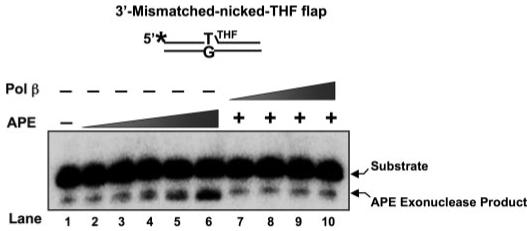
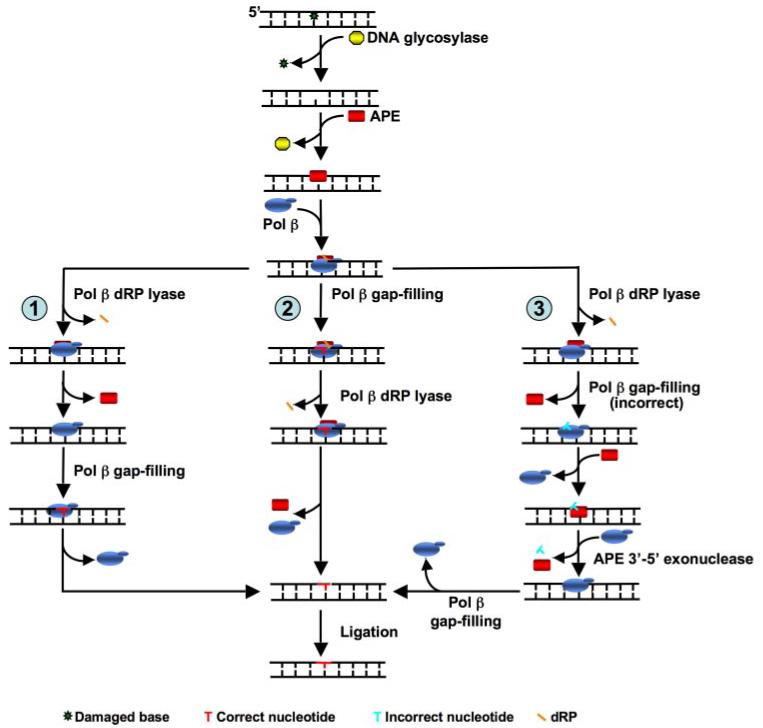
The individual steps in single-nucleotide base excision repair (SN-BER) are coordinated to enable efficient repair without accumulation of cytotoxic DNA intermediates. The DNA transactions and various proteins involved in SN-BER of abasic sites are well known in mammalian systems. Yet, despite a wealth of information on SN-BER, the mechanism of step-by-step coordination is poorly understood. In this study we conducted experiments toward understanding step-by-step coordination during BER by comparing DNA binding specificities of two major human SN-BER enzymes, apurinic/aprymidinic endonuclease 1 (APE) and DNA polymerase beta (Pol beta). It is known that these enzymes do not form a stable complex in solution. For each enzyme, we found that DNA binding specificity appeared sufficient to explain the sequential processing of BER intermediates. In addition, however, we identified at higher enzyme concentrations a ternary complex of APE.Pol beta.DNA that formed specifically at BER intermediates containing a 5'-deoxyribose phosphate group. Formation of this ternary complex was associated with slightly stronger Pol beta gap-filling and much stronger 5'-deoxyribose phosphate lyase activities than was observed with the Pol beta.DNA binary complex. These results indicate that step-by-step coordination in SN-BER can rely on DNA binding specificity inherent in APE and Pol beta, although coordination also may be facilitated by APE.Pol beta.DNA ternary complex formation with appropriate enzyme expression levels or enzyme recruitment to sites of repair.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
