

Epigenetic disruption of ribosomal RNA genes and nucleolar architecture in DNA methyltransferase 1 (Dnmt1) deficient cells
- PMID: 17355984
- PMCID: PMC1874631
- DOI: 10.1093/nar/gkm118
Epigenetic disruption of ribosomal RNA genes and nucleolar architecture in DNA methyltransferase 1 (Dnmt1) deficient cells
Abstract
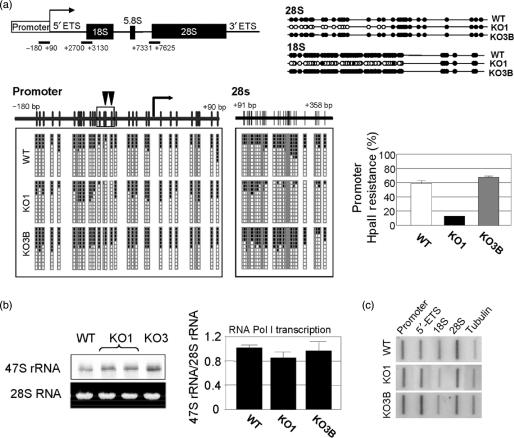
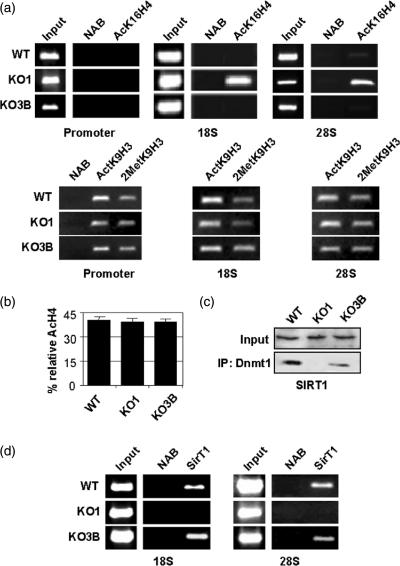
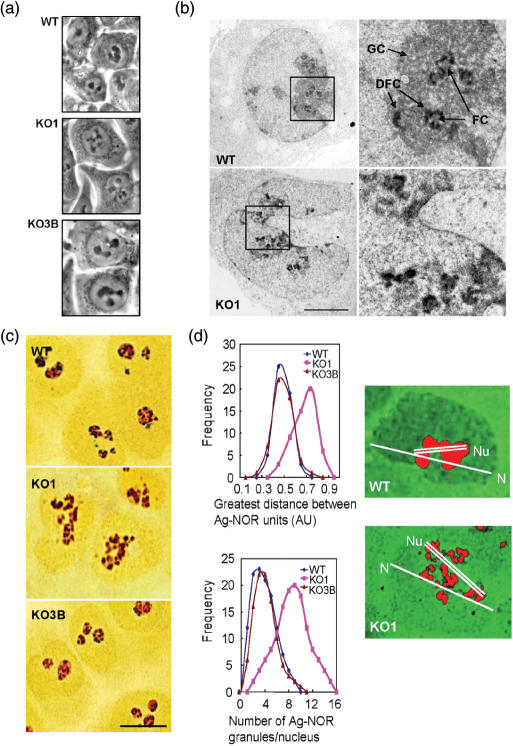
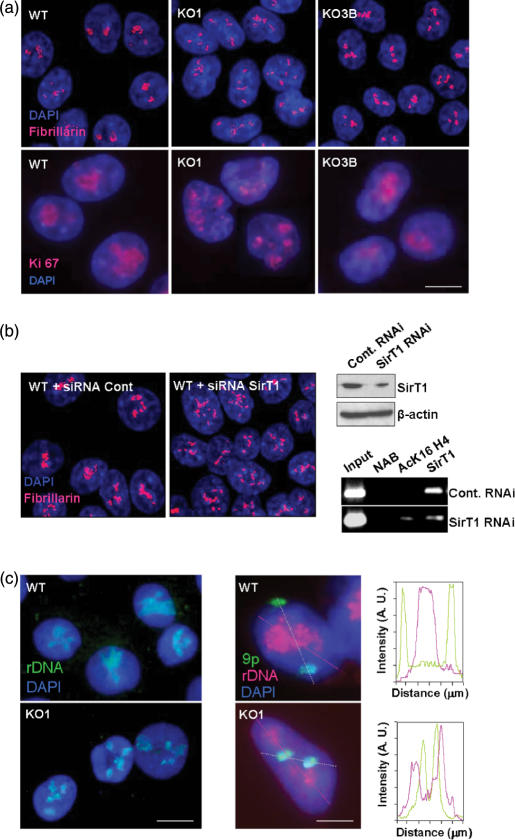
The nucleolus is the site of ribosome synthesis in the nucleus, whose integrity is essential. Epigenetic mechanisms are thought to regulate the activity of the ribosomal RNA (rRNA) gene copies, which are part of the nucleolus. Here we show that human cells lacking DNA methyltransferase 1 (Dnmt1), but not Dnmt33b, have a loss of DNA methylation and an increase in the acetylation level of lysine 16 histone H4 at the rRNA genes. Interestingly, we observed that SirT1, a NAD+-dependent histone deacetylase with a preference for lysine 16 H4, interacts with Dnmt1; and SirT1 recruitment to the rRNA genes is abrogated in Dnmt1 knockout cells. The DNA methylation and chromatin changes at ribosomal DNA observed are associated with a structurally disorganized nucleolus, which is fragmented into small nuclear masses. Prominent nucleolar proteins, such as Fibrillarin and Ki-67, and the rRNA genes are scattered throughout the nucleus in Dnmt1 deficient cells. These findings suggest a role for Dnmt1 as an epigenetic caretaker for the maintenance of nucleolar structure.
Figures
References
-
- Carmo-Fonseca M, Mendes-Soares L, Campos I. To be or not to be in the nucleolus. Nat. Cell Biol. 2000;2:E107–E112. - PubMed
-
- Grummt I, Pikaard CS. Epigenetic silencing of RNA polymerase I transcription. Nat. Rev. Mol. Cell Biol. 2003;4:641–649. - PubMed
-
- McStay B. Nucleolar dominance: a model for rRNA gene silencing. Genes Dev. 2006;20:1207–1214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
