

Retinal pathology and skin barrier defect in mice carrying a Stargardt disease-3 mutation in elongase of very long chain fatty acids-4
- PMID: 17356513
- PMCID: PMC2633486
Retinal pathology and skin barrier defect in mice carrying a Stargardt disease-3 mutation in elongase of very long chain fatty acids-4
Abstract
Purpose: Autosomal dominant Stargardt disease-3 (STGD3) is caused by mutations in elongase of very long chain fatty acids-4 (ELOVL4). The goal of this study was to generate and characterize heterozygous and homozygous knockin-mice that carry a human STGD3 pathogenic mutation in the mouse Elovl4 gene.
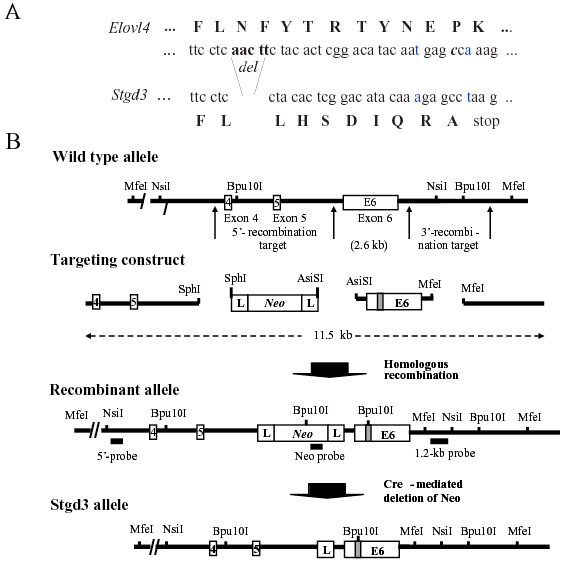
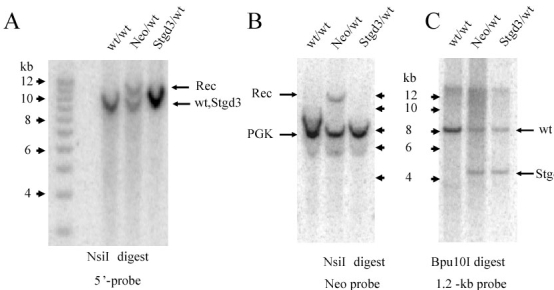
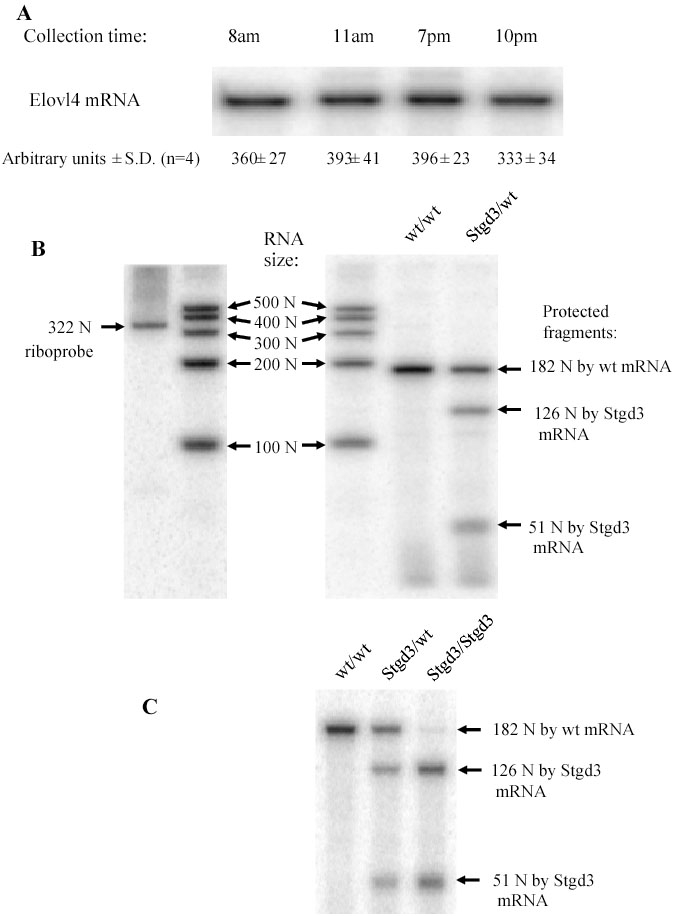
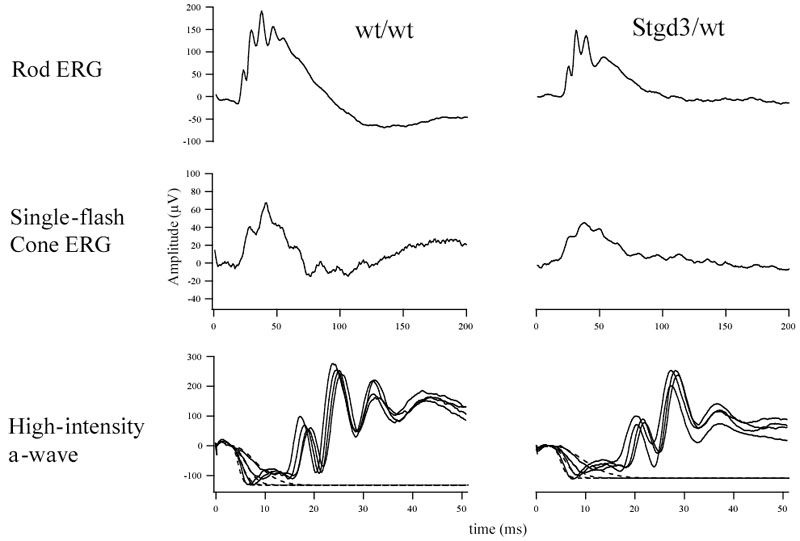
Methods: Recombinant Stgd3-knockin mice were generated using a DNA construct which introduced a pathogenic five-base pair deletion and two point mutations in exon 6 of the Elovl4 gene. Stgd3-mouse genotypes were confirmed by Southern blot analysis and expression of wild-type (wt) and mutated Elovl4 mRNAs assayed by nuclease protection assay. The retinal phenotype of heterozygous Stgd3 mice was characterized by morphological studies, elecroretinographic (ERG) analysis and assay of lipofuscin accumulation. Homozygous Stgd3 mice were examined for both retinal and gross morphology. They were also analyzed for skin morphology and skin barrier function, and for epidermal lipid content using high performance liquid chromatography (HPLC) combined with mass spectrometry (MS).
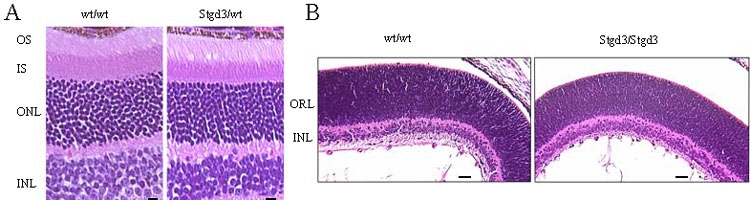
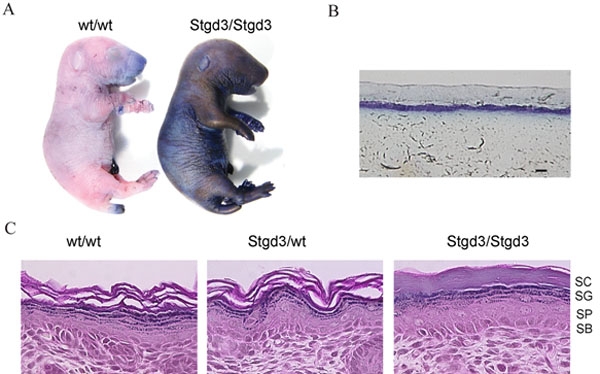
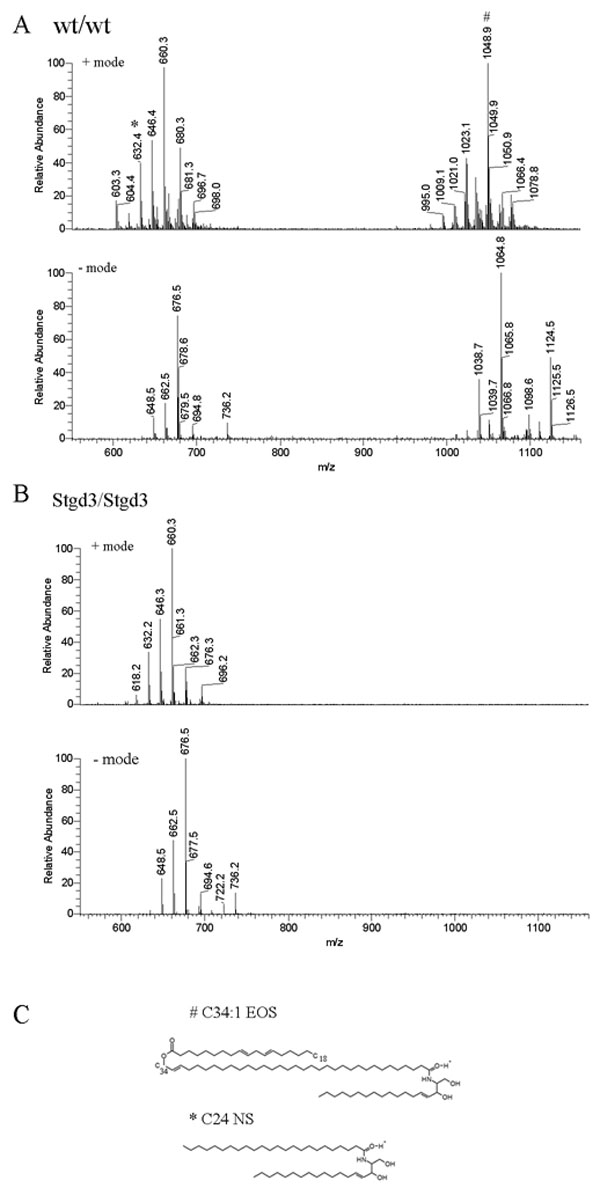
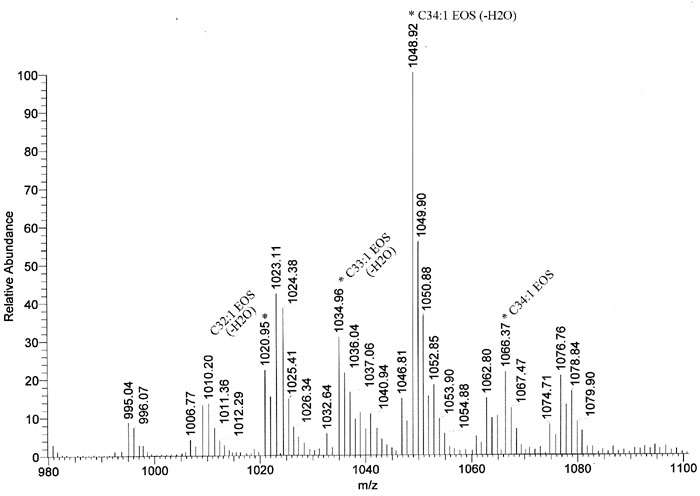
Results: The Stgd3 allele codes for a truncated mouse Elovl4 protein, which also contains the same aberrant 8-amino acid C-terminus encoded by the human pathogenic STGD3 allele. Heterozygous Stgd3 mice expressed equal amounts of both wt and mutant Elovl4 mRNAs in the retina, showed no significant changes in retinal morphology, but did show accumulation of lipofuscin and reduced visual function. Homozygous Stgd3 mice were born with an expected Mendelian frequency, without any initial gross anatomical or behavioral abnormalities. By 6-12 h postpartum, they became dehydrated and died. A skin permeability assay detected a defect in epidermal barrier function. Homozygous mutant epidermis expressed a normal content of mutated Elovl4 mRNA and contained all four epidermal cellular layers. HPLC/MS analysis of epidermal lipids revealed the presence of all barrier lipids with the exception of the complete absence of acylceramides, the critical lipids for barrier function of the skin.
Conclusions: The generated Stgd3-knockin mice are a genetic model of human STGD3 and reproduce features of the human disease: accumulation of lipofuscin and reduced visual functions. Homozygous Stgd3 mice showed a complete absence of acylceramides from the epidermis. Their absence suggests a role for Elovl4 in acylceramide synthesis, and in particular, a role in the synthesis of the unique very long chain C30-C40 fatty acids present in skin acylceramides.
Figures

References
-
- Zhang K, Kniazeva M, Han M, Li W, Yu Z, Yang Z, Li Y, Metzker ML, Allikmets R, Zack DJ, Kakuk LE, Lagali PS, Wong PW, MacDonald IM, Sieving PA, Figueroa DJ, Austin CP, Gould RJ, Ayyagari R, Petrukhin K. A 5-bp deletion in ELOVL4 is associated with two related forms of autosomal dominant macular dystrophy. Nat Genet. 2001;27:89–93. - PubMed
-
- Edwards AO, Donoso LA, Ritter R., 3rd A novel gene for autosomal dominant Stargardt-like macular dystrophy with homology to the SUR4 protein family. Invest Ophthalmol Vis Sci. 2001;42:2652–63. - PubMed
-
- Bernstein PS, Tammur J, Singh N, Hutchinson A, Dixon M, Pappas CM, Zabriskie NA, Zhang K, Petrukhin K, Leppert M, Allikmets R. Diverse macular dystrophy phenotype caused by a novel complex mutation in the ELOVL4 gene. Invest Ophthalmol Vis Sci. 2001;42:3331–6. - PubMed
-
- Maugeri A, Meire F, Hoyng CB, Vink C, Van Regemorter N, Karan G, Yang Z, Cremers FP, Zhang K. A novel mutation in the ELOVL4 gene causes autosomal dominant Stargardt-like macular dystrophy. Invest Ophthalmol Vis Sci. 2004;45:4263–7. - PubMed
-
- Stone EM, Nichols BE, Kimura AE, Weingeist TA, Drack A, Sheffield VC. Clinical features of a Stargardt-like dominant progressive macular dystrophy with genetic linkage to chromosome 6q. Arch Ophthalmol. 1994;112:765–72. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases