

Detyrosinated microtubule protrusions in suspended mammary epithelial cells promote reattachment
- PMID: 17359970
- PMCID: PMC3132414
- DOI: 10.1016/j.yexcr.2007.02.001
Detyrosinated microtubule protrusions in suspended mammary epithelial cells promote reattachment
Abstract
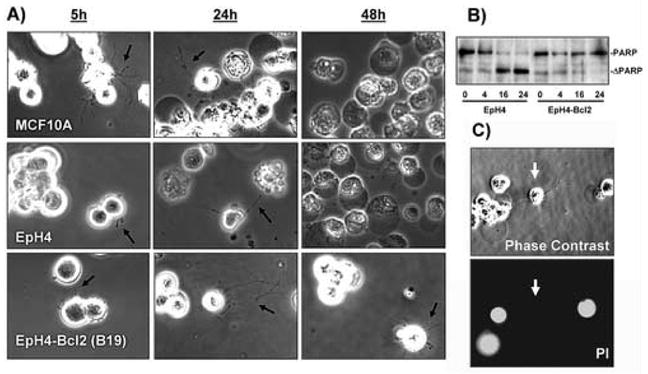
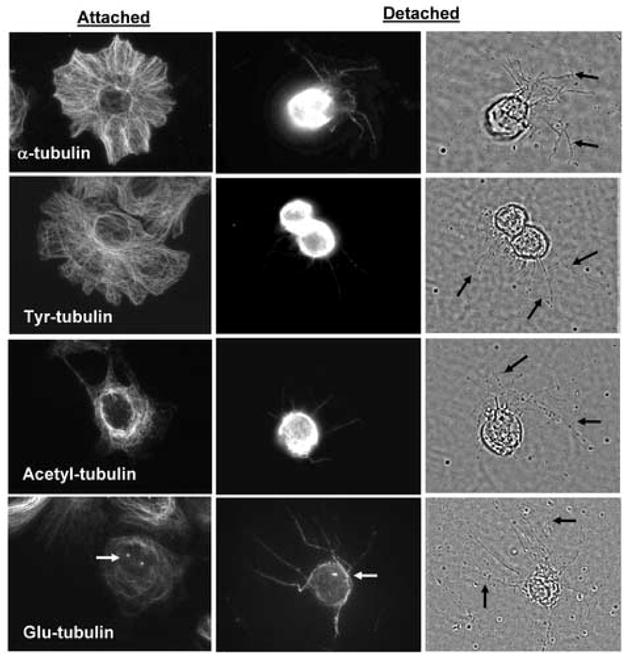
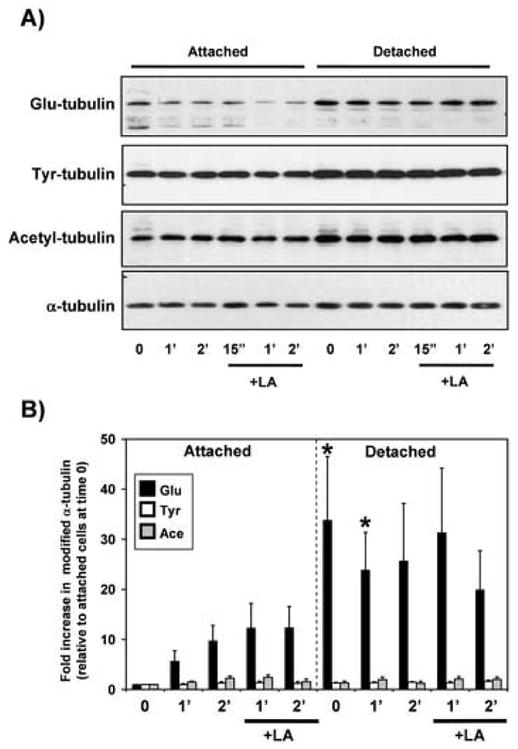
Breast tumor cells enter the bloodstream long before the development of clinically evident metastasis. However, the early presence of such bloodborne cells predicts poor patient outcome. Nearly 90% of human breast tumors arise as carcinomas from mammary epithelial cells, so it is important to study how these cells respond to the detached conditions that they would experience in the bloodstream. We report here that mammary epithelial cell lines produce long and dynamic protrusions of the plasma membrane when detached. Although human and mouse mammary epithelial cell lines die by apoptosis within 16 h of detachment, this protrusive response persists for days in cells overexpressing either Bcl-2 or Bcl-xL. Unlike actin-dependent invadopodia and podosomes, these protrusions are actually enhanced by actin depolymerization with Cytochalasin-D or Latrunculin-A. Immunofluorescence and Western blotting demonstrate that the protrusions are enriched in detyrosinated Glu-tubulin, a post-translationally modified form of alpha-tubulin that is found in stabilized microtubules. Video microscopy indicates that these protrusions promote cell-cell attachment, and inhibiting microtubule-based protrusions correlates with reduced extracellular matrix attachment. Since bloodborne metastasis depends on both cell-cell and cell-matrix attachment, microtubule-based protrusions in detached mammary epithelial cells provide a novel mechanism that could influence the metastatic spread of breast tumors.
Figures



References
-
- Naumov GN, MacDonald IC, Weinmeister PM, Kerkvliet N, Nadkarni KV, Wilson SM, Morris VL, Groom AC, Chambers AF. Persistence of solitary mammary carcinoma cells in a secondary site: a possible contributor to dormancy. Cancer Res. 2002;62:2162–8. - PubMed
-
- Naumov GN, MacDonald IC, Chambers AF, Groom AC. Solitary cancer cells as a possible source of tumour dormancy? Semin Cancer Biol. 2001;11:271–6. - PubMed
-
- Schmidt-Kittler O, Ragg T, Daskalakis A, Granzow M, Ahr A, Blankenstein TJ, Kaufmann M, Diebold J, Arnholdt H, Muller P, Bischoff J, Harich D, Schlimok G, Riethmuller G, Eils R, Klein CA. From latent disseminated cells to overt metastasis: genetic analysis of systemic breast cancer progression. Proc Natl Acad Sci U S A. 2003;100:7737–42. - PMC - PubMed
-
- Chambers AF, Groom AC, MacDonald IC. Dissemination and growth of cancer cells in metastatic sites. Nat Rev Cancer. 2002;2:563–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
