

Intersubunit movement is required for ribosomal translocation
- PMID: 17360328
- PMCID: PMC1829233
- DOI: 10.1073/pnas.0700762104
Intersubunit movement is required for ribosomal translocation
Abstract
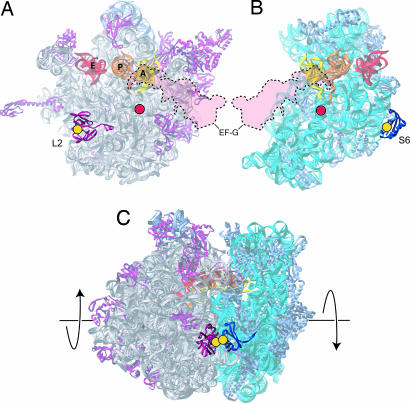
Translocation of tRNA and mRNA during protein synthesis is believed to be coupled to structural changes in the ribosome. The "ratchet model," based on cryo-EM reconstructions of ribosome complexes, invokes relative movement of the 30S and 50S ribosomal subunits in this process; however, evidence that directly demonstrates a requirement for intersubunit movement during translocation is lacking. To address this problem, we created an intersubunit disulfide cross-link to restrict potential movement. The cross-linked ribosomes were unable to carry out polypeptide synthesis; this inhibition was completely reversed upon reduction of the disulfide bridge. In vitro assays showed that the cross-linked ribosomes were specifically blocked in elongation factor G-dependent translocation. These findings show that intersubunit movement is required for ribosomal translocation, accounting for the universal two-subunit architecture of ribosomes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Conformational changes of the small ribosomal subunit during elongation factor G-dependent tRNA-mRNA translocation.J Mol Biol. 2004 Nov 5;343(5):1183-94. doi: 10.1016/j.jmb.2004.08.097. J Mol Biol. 2004. PMID: 15491605
-
Spontaneous intersubunit rotation in single ribosomes.Mol Cell. 2008 Jun 6;30(5):578-88. doi: 10.1016/j.molcel.2008.05.004. Mol Cell. 2008. PMID: 18538656 Free PMC article.
-
On the pathway of ribosomal translocation.Int J Biol Macromol. 2016 Nov;92:401-415. doi: 10.1016/j.ijbiomac.2016.07.048. Epub 2016 Jul 16. Int J Biol Macromol. 2016. PMID: 27431796
-
Structural insights into ribosome translocation.Wiley Interdiscip Rev RNA. 2016 Sep;7(5):620-36. doi: 10.1002/wrna.1354. Epub 2016 Apr 27. Wiley Interdiscip Rev RNA. 2016. PMID: 27117863 Free PMC article. Review.
-
Synchronous tRNA movements during translocation on the ribosome are orchestrated by elongation factor G and GTP hydrolysis.Bioessays. 2014 Oct;36(10):908-18. doi: 10.1002/bies.201400076. Epub 2014 Aug 13. Bioessays. 2014. PMID: 25118068 Review.
Cited by
-
Coordinated conformational and compositional dynamics drive ribosome translocation.Nat Struct Mol Biol. 2013 Jun;20(6):718-27. doi: 10.1038/nsmb.2567. Epub 2013 Apr 28. Nat Struct Mol Biol. 2013. PMID: 23624862 Free PMC article.
-
Translocation as continuous movement through the ribosome.RNA Biol. 2016 Dec;13(12):1197-1203. doi: 10.1080/15476286.2016.1240140. Epub 2016 Nov 1. RNA Biol. 2016. PMID: 27801619 Free PMC article.
-
Time-resolved cryo-EM visualizes ribosomal translocation with EF-G and GTP.Nat Commun. 2021 Dec 13;12(1):7236. doi: 10.1038/s41467-021-27415-0. Nat Commun. 2021. PMID: 34903725 Free PMC article.
-
Taura syndrome virus IRES initiates translation by binding its tRNA-mRNA-like structural element in the ribosomal decoding center.Proc Natl Acad Sci U S A. 2014 Jun 24;111(25):9139-44. doi: 10.1073/pnas.1406335111. Epub 2014 Jun 9. Proc Natl Acad Sci U S A. 2014. PMID: 24927574 Free PMC article.
-
The ribosome moves: RNA mechanics and translocation.Nat Struct Mol Biol. 2017 Dec 7;24(12):1021-1027. doi: 10.1038/nsmb.3505. Nat Struct Mol Biol. 2017. PMID: 29215639 Free PMC article.
References
-
- Tissiéres A, Watson JD. Nature. 1958;182:778–780. - PubMed
-
- Spirin AS. Dokl Akad Nauk SSSR. 1968;179:1467–1470. - PubMed
-
- Bretscher MS. Nature. 1968;218:675–677. - PubMed
-
- Moazed D, Noller HF. Nature. 1989;342:142–148. - PubMed
-
- Noller HF, Moazed D, Stern S, Powers T, Allen PA, Robertson JM, Weiser B, Triman K. In: The Ribosome Structure, Function, and Evolution. Hill WE, Dahlberf A, Garrett RA, Moore PB, Schlessinger D, Warner JR, editors. Washington, DC: Am Soc Microbiol; 1990. pp. 73–92.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
