

Reduction of 1-Cys peroxiredoxins by ascorbate changes the thiol-specific antioxidant paradigm, revealing another function of vitamin C
- PMID: 17360337
- PMCID: PMC1829234
- DOI: 10.1073/pnas.0700481104
Reduction of 1-Cys peroxiredoxins by ascorbate changes the thiol-specific antioxidant paradigm, revealing another function of vitamin C
Abstract
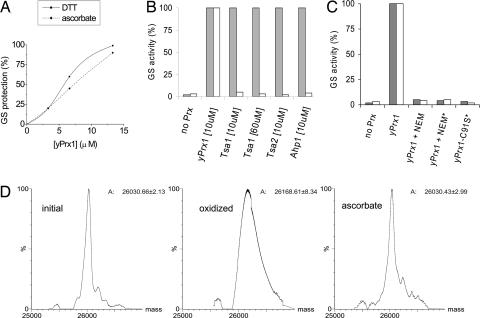
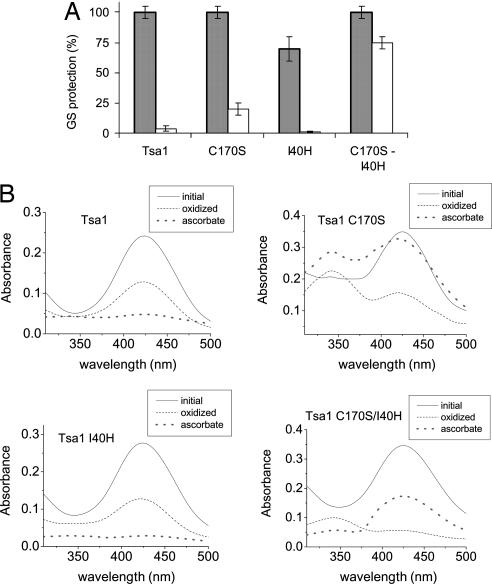
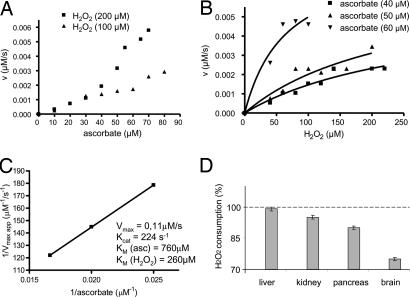
Peroxiredoxins (Prx) are widely distributed peroxidases that can be divided into 1-Cys and 2-Cys Prx groups based on the number of conserved cysteine residues that participate in their catalytical cycle. Prx have been described to be strictly dependent on thiols, but here, we show that ascorbate (vitamin C) also reduces 1-Cys Prx, but not 2-Cys Prx, from several taxonomic groups. Reduction by ascorbate is partly related to the fact that the oxidized form of 1-Cys Prx is a stable sulfenic acid (Cys-SOH) instead of a disulfide. In addition, a histidine residue in the active site is required. In fact, we engineered a 2-Cys Prx with these two features, and it displayed ascorbate peroxidase activity. These data represent a breakthrough in the thiol-specific antioxidant paradigm. Ascorbate may be the long-sought-after biological reductant of 1-Cys Prx. Because ascorbate is present in high amounts in cells, the ascorbate/protein sulfenic acid pair represents an aspect of redox biochemistry that has yet to be explored in vivo.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Imlay JA. Annu Rev Microbiol. 2003;57:395–418. - PubMed
-
- Rumsey SC, Levine M. J Nutr Biochem. 1998;9:116–130.
-
- Wood ZA, Schroder E, Harris JR, Poole LB. Trends Biochem Sci. 2003;28:32–40. - PubMed
-
- Neumann CA, Krause DS, Carman CV, Das S, Dubey DP, Abraham JL, Bronson RT, Fujiwara Y, Orkin SH, Van Etten RA. Nature. 2003;424:561–565. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
