

Lipidomics reveals control of Mycobacterium tuberculosis virulence lipids via metabolic coupling
- PMID: 17360366
- PMCID: PMC1829275
- DOI: 10.1073/pnas.0610634104
Lipidomics reveals control of Mycobacterium tuberculosis virulence lipids via metabolic coupling
Abstract
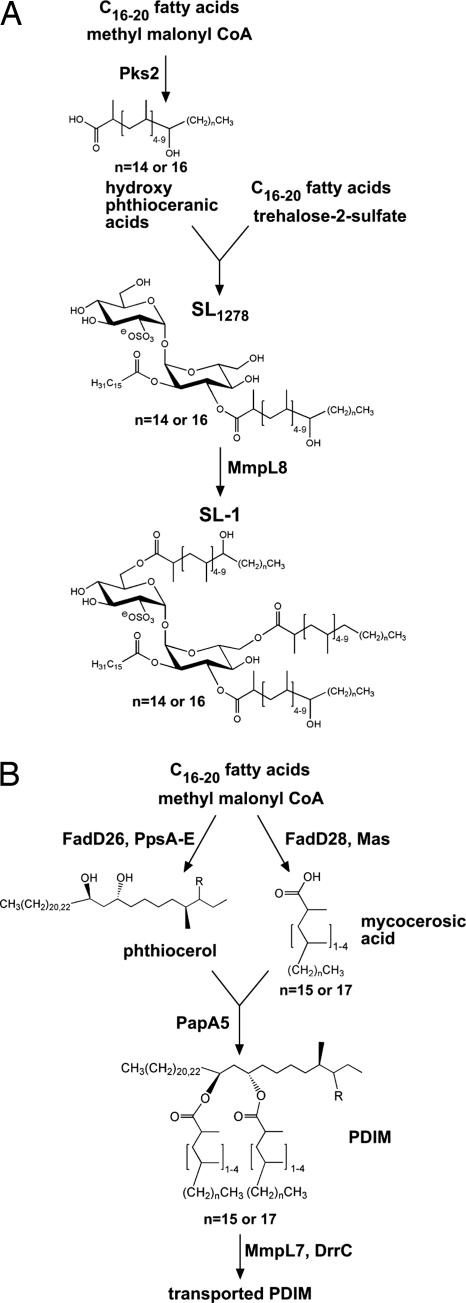
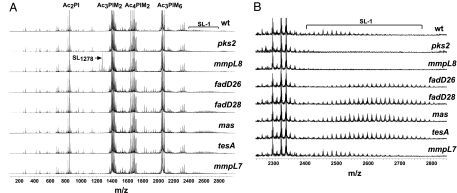
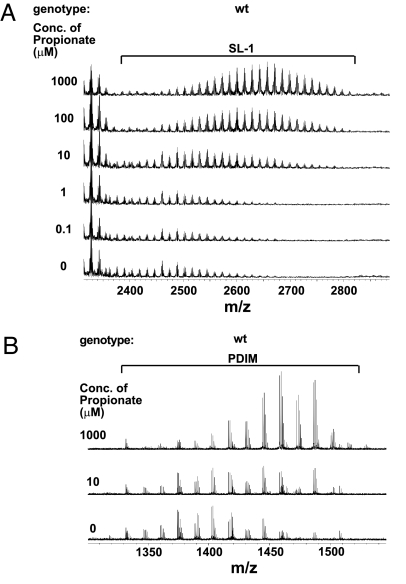
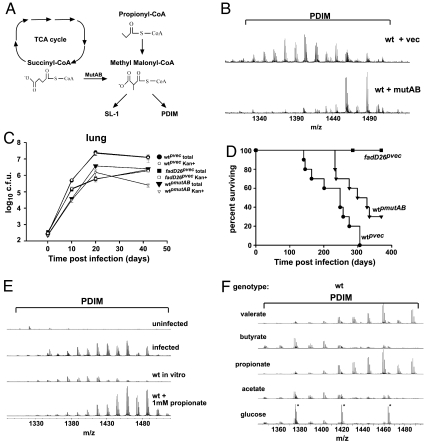
Mycobacterium tuberculosis synthesizes specific polyketide lipids that interact with the host and are required for virulence. Using a mass spectrometric approach to simultaneously monitor hundreds of lipids, we discovered that the size and abundance of two lipid virulence factors, phthiocerol dimycocerosate (PDIM) and sulfolipid-1 (SL-1), are controlled by the availability of a common precursor, methyl malonyl CoA (MMCoA). Consistent with this view, increased levels of MMCoA led to increased abundance and mass of both PDIM and SL-1. Furthermore, perturbation of MMCoA metabolism attenuated pathogen replication in mice. Importantly, we detected increased PDIM synthesis in bacteria growing within host tissues and in bacteria grown in culture on odd-chain fatty acids. Because M. tuberculosis catabolizes host lipids to grow during infection, we propose that growth of M. tuberculosis on fatty acids in vivo leads to increased flux of MMCoA through lipid biosynthetic pathways, resulting in increased virulence lipid synthesis. Our results suggest that the shift to host lipid catabolism during infection allows for increased virulence lipid anabolism by the bacterium.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Brennan PJ, Nikaido H. Annu Rev Biochem. 1995;64:29–63. - PubMed
-
- Cox JS, Chen B, McNeil M, Jacobs WR., Jr Nature. 1999;402:79–83. - PubMed
-
- Camacho LR, Ensergueix D, Perez E, Gicquel B, Guilhot C. Mol Microbiol. 1999;34:257–267. - PubMed
-
- Domenech P, Reed MB, Dowd CS, Manca C, Kaplan G, Barry CE., III J Biol Chem. 2004;279:21257–21265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
