

Regulation of poly(ADP-ribose) polymerase 1 activity by the phosphorylation state of the nuclear NAD biosynthetic enzyme NMN adenylyl transferase 1
- PMID: 17360427
- PMCID: PMC1820658
- DOI: 10.1073/pnas.0609211104
Regulation of poly(ADP-ribose) polymerase 1 activity by the phosphorylation state of the nuclear NAD biosynthetic enzyme NMN adenylyl transferase 1
Abstract
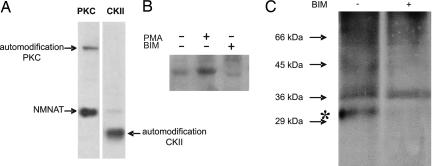
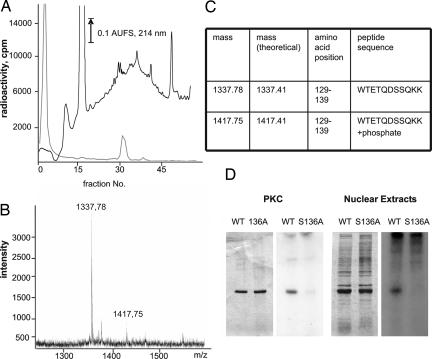
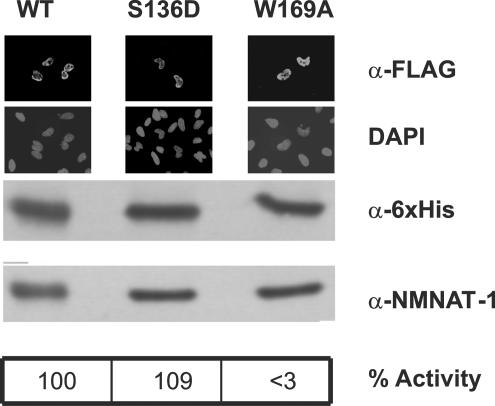
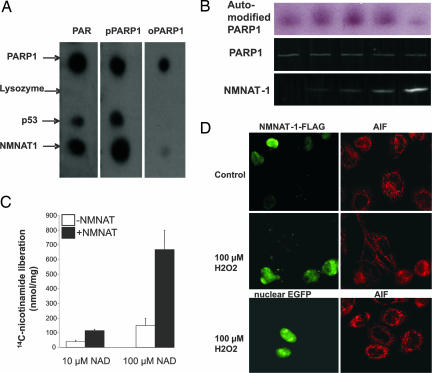
Nuclear NAD(+) metabolism constitutes a major component of signaling pathways. It includes NAD(+)-dependent protein deacetylation by members of the Sir2 family and protein modification by poly(ADP-ribose) polymerase 1 (PARP-1). PARP-1 has emerged as an important mediator of processes involving DNA rearrangements. High-affinity binding to breaks in DNA activates PARP-1, which attaches poly(ADP-ribose) (PAR) to target proteins. NMN adenylyl transferases (NMNATs) catalyze the final step of NAD(+) biosynthesis. We report here that the nuclear isoform NMNAT-1 stimulates PARP-1 activity and binds to PAR. Its overexpression in HeLa cells promotes the relocation of apoptosis-inducing factor from the mitochondria to the nucleus, a process known to depend on poly(ADP-ribosyl)ation. Moreover, NMNAT-1 is subject to phosphorylation by protein kinase C, resulting in reduced binding to PAR. Mimicking phosphorylation, substitution of the target serine residue by aspartate precludes PAR binding and stimulation of PARP-1. We conclude that, depending on its state of phosphorylation, NMNAT-1 binds to activated, automodifying PARP-1 and thereby amplifies poly(ADP-ribosyl)ation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Magni G, Amici A, Emanuelli M, Raffaelli N, Ruggieri S. Adv Enzymol Relat Areas Mol Biol. 1999;73:135–182. - PubMed
-
- Berger F, Lau C, Dahlmann M, Ziegler M. J Biol Chem. 2005;280:36334–36341. - PubMed
-
- Schweiger M, Hennig K, Lerner F, Niere M, Hirsch-Kauffmann M, Specht T, Weise C, Oei SL, Ziegler M. FEBS Lett. 2001;492:95–100. - PubMed
-
- Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Cohen H, Lin SS, Manchester JK, Gordon JI, Sinclair DA. J Biol Chem. 2002;277:18881–18890. - PubMed
-
- Berger F, Ramirez-Hernandez MH, Ziegler M. Trends Biochem Sci. 2004;29:111–118. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
