

Cortical area size dictates performance at modality-specific behaviors
- PMID: 17360492
- PMCID: PMC1820724
- DOI: 10.1073/pnas.0611723104
Cortical area size dictates performance at modality-specific behaviors
Abstract
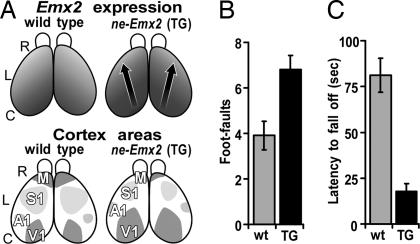
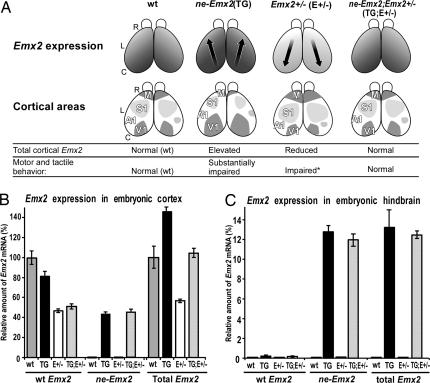
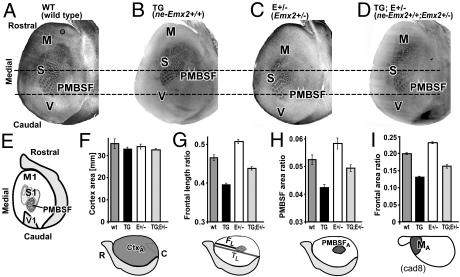
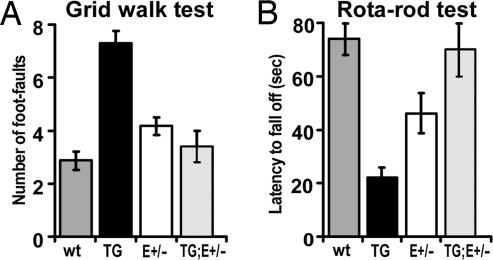
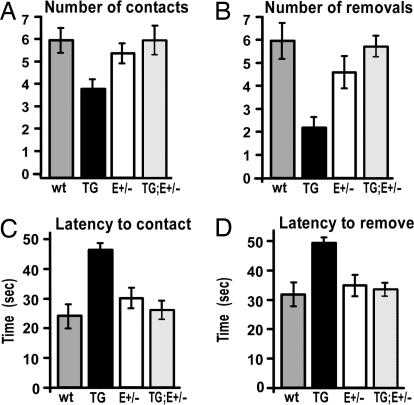
The mammalian neocortex is organized into unique areas that serve functions such as sensory perception and modality-specific behaviors. The sizes of primary cortical areas vary across species, and also within a species, raising the question of whether area size dictates behavioral performance. We show that adult mice genetically engineered to overexpress the transcription factor EMX2 in embryonic cortical progenitor cells, resulting in reductions in sizes of somatosensory and motor areas, exhibit significant deficiencies at tactile and motor behaviors. Even increasing the size of sensorimotor areas by decreasing cortical EMX2 levels can lead to diminished sensorimotor behaviors. Genetic crosses that retain ectopic Emx2 transgene expression subcortically but restore cortical Emx2 expression to wild-type levels also restore cortical areas to wild-type sizes and in parallel restore tactile and motor behaviors to wild-type performance. These findings show that area size can dictate performance at modality-specific behaviors and suggest that areas have an optimal size, influenced by parameters of its neural system, for maximum behavioral performance. This study underscores the importance of establishing during embryonic development appropriate levels of regulatory proteins that determine area sizes, thereby influencing behavior later in life.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Brodmann K. Allg Z Psychiatr (Berlin) 1918;74:564–568.
-
- Dougherty RF, Koch VM, Brewer AA, Fischer B, Modersitzki J, Wandell BA. J Vis. 2003;3:586–598. - PubMed
-
- Stensaas SS, Eddington DK, Dobelle WH. J Neurosurg. 1974;40:747–755. - PubMed
-
- White LE, Andrews TJ, Hulette C, Richards A, Groelle M, Paydarfar J, Purves D. Cereb Cortex. 1997;7:31–47. - PubMed
-
- White LE, Andrews TJ, Hulette C, Richards A, Groelle M, Paydarfar J, Purves D. Cereb Cortex. 1997;7:18–30. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
