

Monitoring the function of membrane transport proteins in detergent-solubilized form
- PMID: 17360689
- PMCID: PMC1805550
- DOI: 10.1073/pnas.0609573104
Monitoring the function of membrane transport proteins in detergent-solubilized form
Abstract
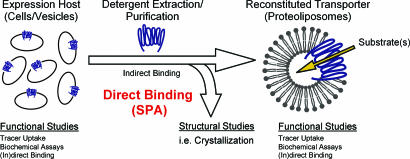
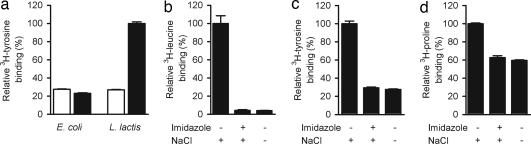
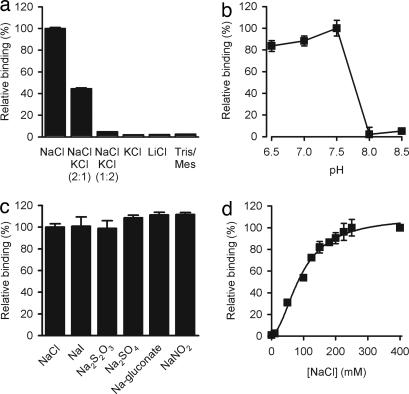
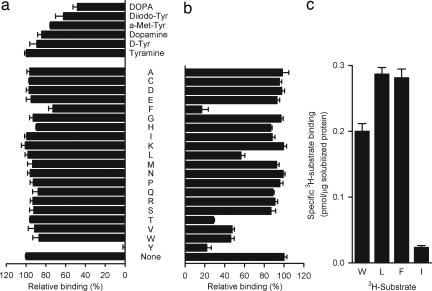
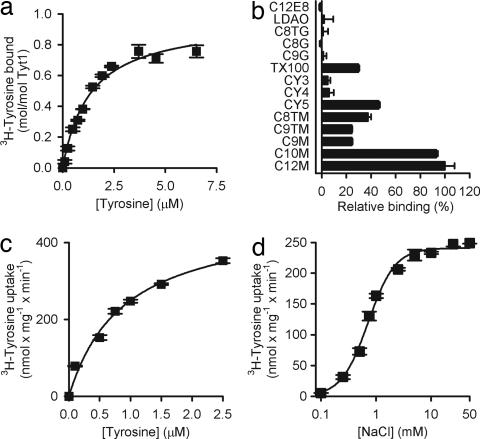
Transport proteins constitute approximately 10% of most proteomes and play vital roles in the translocation of solutes across membranes of all organisms. Their (dys)function is implicated in many disorders, making them frequent targets for pharmacotherapy. The identification of substrates for members of this large protein family, still replete with many orphans of unknown function, has proven difficult, in part because high-throughput screening is greatly complicated by endogenous transporters present in many expression systems. In addition, direct structural studies require that transporters be extracted from the membrane with detergent, thereby precluding transport measurements because of the lack of a vectorial environment and necessitating reconstitution into proteoliposomes for activity measurements. Here, we describe a direct scintillation proximity-based radioligand-binding assay for determining transport protein function in crude cell extracts and in purified form. This rapid and universally applicable assay with advantages over cell-based platforms will greatly facilitate the identification of substrates for many orphan transporters and allows monitoring the function of transport proteins in a nonmembranous environment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
