

Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots
- PMID: 17360695
- PMCID: PMC1802001
- DOI: 10.1073/pnas.0607703104
Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots
Abstract
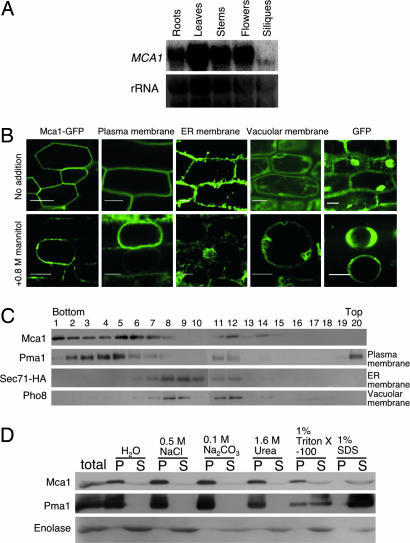
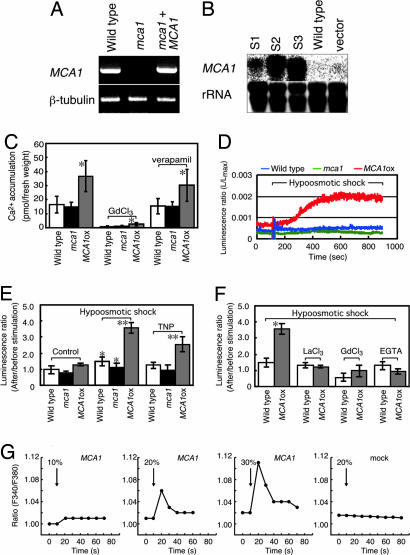
Plants can sense and respond to mechanical stimuli, like animals. An early mechanism of mechanosensing and response is speculated to be governed by as-yet-unidentified sensory complexes containing a Ca(2+)-permeable, stretch-activated (SA) channel. However, the components or regulators of such complexes are poorly understood at the molecular level in plants. Here, we report the molecular identification of a plasma membrane protein (designated Mca1) that correlates Ca(2+) influx with mechanosensing in Arabidopsis thaliana. MCA1 cDNA was cloned by the functional complementation of lethality of a yeast mid1 mutant lacking a putative Ca(2+)-permeable SA channel component. Mca1 was localized to the yeast plasma membrane as an integral membrane protein and mediated Ca(2+) influx. Mca1 also increased [Ca(2+)](cyt) upon plasma membrane distortion in Arabidopsis. The growth of MCA1-overexpressing plants was impaired in a high-calcium but not a low-calcium medium. The primary roots of mca1-null plants failed to penetrate a harder agar medium from a softer one. These observations demonstrate that Mca1 plays a crucial role in a Ca(2+)-permeable SA channel system that leads to mechanosensing in Arabidopsis. We anticipate our findings to be a starting point for a deeper understanding of the molecular mechanisms of mechanotransduction in plants.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis.Plant Physiol. 2010 Mar;152(3):1284-96. doi: 10.1104/pp.109.147371. Epub 2010 Jan 22. Plant Physiol. 2010. PMID: 20097794 Free PMC article.
-
Transmembrane Topologies of Ca2+-permeable Mechanosensitive Channels MCA1 and MCA2 in Arabidopsis thaliana.J Biol Chem. 2015 Dec 25;290(52):30901-9. doi: 10.1074/jbc.M115.692574. Epub 2015 Nov 10. J Biol Chem. 2015. PMID: 26555262 Free PMC article.
-
Ca2+-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca2+ increase and cold tolerance in Arabidopsis.Sci Rep. 2018 Jan 11;8(1):550. doi: 10.1038/s41598-017-17483-y. Sci Rep. 2018. PMID: 29323146 Free PMC article.
-
Physiological roles of Arabidopsis MCA1 and MCA2 based on their dynamic expression patterns.J Plant Res. 2024 Sep;137(5):785-797. doi: 10.1007/s10265-024-01575-8. Epub 2024 Aug 28. J Plant Res. 2024. PMID: 39196431 Free PMC article. Review.
-
New candidates for mechano-sensitive channels potentially involved in gravity sensing in Arabidopsis thaliana.Plant Biol (Stuttg). 2014 Jan;16 Suppl 1:39-42. doi: 10.1111/plb.12044. Epub 2013 Jun 4. Plant Biol (Stuttg). 2014. PMID: 23731064 Review.
Cited by
-
Plant cell wall integrity maintenance as an essential component of biotic stress response mechanisms.Front Plant Sci. 2012 Apr 23;3:77. doi: 10.3389/fpls.2012.00077. eCollection 2012. Front Plant Sci. 2012. PMID: 22629279 Free PMC article.
-
VIP1 is very important/interesting protein 1 regulating touch responses of Arabidopsis.Plant Signal Behav. 2016 Jun 2;11(6):e1187358. doi: 10.1080/15592324.2016.1187358. Plant Signal Behav. 2016. PMID: 27171129 Free PMC article.
-
ROOT PENETRATION INDEX 3, a major quantitative trait locus associated with root system penetrability in Arabidopsis.J Exp Bot. 2022 Aug 11;73(14):4716-4732. doi: 10.1093/jxb/erac188. J Exp Bot. 2022. PMID: 35512438 Free PMC article.
-
Rare earth elements induce cytoskeleton-dependent and PI4P-associated rearrangement of SYT1/SYT5 endoplasmic reticulum-plasma membrane contact site complexes in Arabidopsis.J Exp Bot. 2020 Jul 6;71(14):3986-3998. doi: 10.1093/jxb/eraa138. J Exp Bot. 2020. PMID: 32179893 Free PMC article.
-
Root osmotic sensing from local perception to systemic responses.Stress Biol. 2022 Sep 5;2(1):36. doi: 10.1007/s44154-022-00054-1. Stress Biol. 2022. PMID: 37676549 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
