

Old World and clade C New World arenaviruses mimic the molecular mechanism of receptor recognition used by alpha-dystroglycan's host-derived ligands
- PMID: 17360738
- PMCID: PMC1900301
- DOI: 10.1128/JVI.02574-06
Old World and clade C New World arenaviruses mimic the molecular mechanism of receptor recognition used by alpha-dystroglycan's host-derived ligands
Abstract
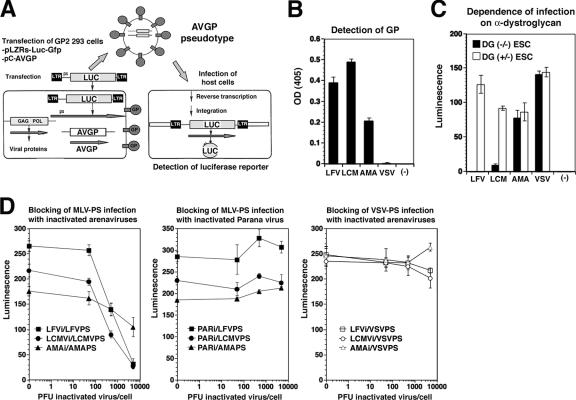
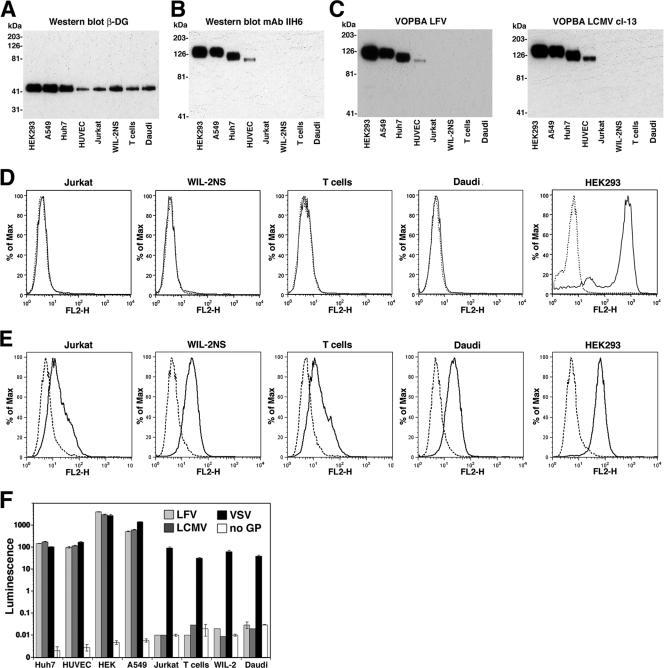
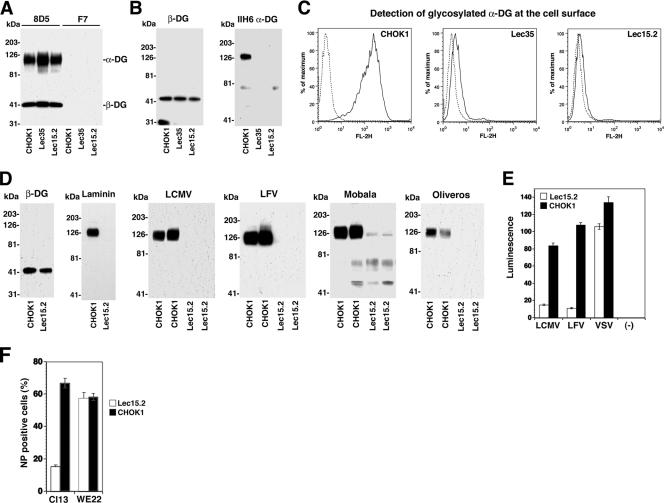
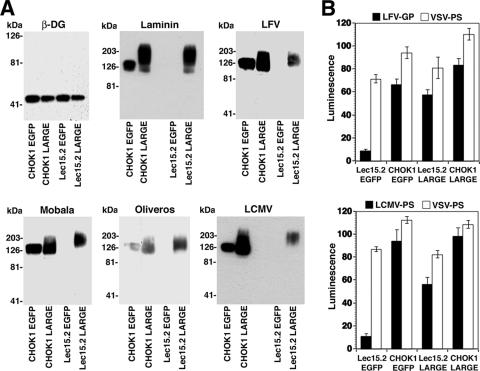
alpha-Dystroglycan (DG) is an important cellular receptor for extracellular matrix (ECM) proteins and also serves as the receptor for Old World arenaviruses Lassa fever virus (LFV) and lymphocytic choriomeningitis virus (LCMV) and clade C New World arenaviruses. In the host cell, alpha-DG is subject to a remarkably complex pattern of O glycosylation that is crucial for its interactions with ECM proteins. Two of these unusual sugar modifications, protein O mannosylation and glycan modifications involving the putative glycosyltransferase LARGE, have recently been implicated in arenavirus binding. Considering the complexity of alpha-DG O glycosylation, our present study was aimed at the identification of the specific O-linked glycans on alpha-DG that are recognized by arenaviruses. As previously shown for LCMV, we found that protein O mannosylation of alpha-DG is crucial for the binding of arenaviruses of distinct phylogenetic origins, including LFV, Mobala virus, and clade C New World arenaviruses. In contrast to the highly conserved requirement for O mannosylation, more generic O glycans present on alpha-DG are dispensable for arenavirus binding. Despite the critical role of O-mannosyl glycans for arenavirus binding under normal conditions, the overexpression of LARGE in cells deficient in O mannosylation resulted in highly glycosylated alpha-DG that was functional as a receptor for arenaviruses. Thus, modifications by LARGE but not O-mannosyl glycans themselves are most likely the crucial structures recognized by arenaviruses. Together, the data demonstrate that arenaviruses recognize the same highly conserved O-glycan structures on alpha-DG involved in ECM protein binding, indicating a strikingly similar mechanism of receptor recognition by pathogen- and host-derived ligands.
Figures
References
-
- Ahmed, R., A. Salmi, L. D. Butler, J. M. Chiller, and M. B. Oldstone. 1984. Selection of genetic variants of lymphocytic choriomeningitis virus in spleens of persistently infected mice. Role in suppression of cytotoxic T lymphocyte response and viral persistence. J. Exp. Med. 160:521-540. - PMC - PubMed
-
- Andac, Z., T. Sasaki, K. Mann, A. Brancaccio, R. Deutzmann, and R. Timpl. 1999. Analysis of heparin, alpha-dystroglycan and sulfatide binding to the G domain of the laminin alpha1 chain by site-directed mutagenesis. J. Mol. Biol. 287:253-264. - PubMed
-
- Barresi, R., and K. P. Campbell. 2006. Dystroglycan: from biosynthesis to pathogenesis of human disease. J. Cell Sci. 119:199-207. - PubMed
-
- Barresi, R., D. E. Michele, M. Kanagawa, H. A. Harper, S. A. Dovicio, J. S. Satz, S. A. Moore, W. Zhang, H. Schachter, J. P. Dumanski, and K. P. Campbell. 2004. LARGE can functionally bypass alpha-dystroglycan glycosylation defects in distinct congenital muscular dystrophy. Nat. Med. 7:696-703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
