

Brain-derived neurotrophic factor promotes long-term potentiation-related cytoskeletal changes in adult hippocampus
- PMID: 17360925
- PMCID: PMC6672589
- DOI: 10.1523/JNEUROSCI.4037-06.2007
Brain-derived neurotrophic factor promotes long-term potentiation-related cytoskeletal changes in adult hippocampus
Abstract
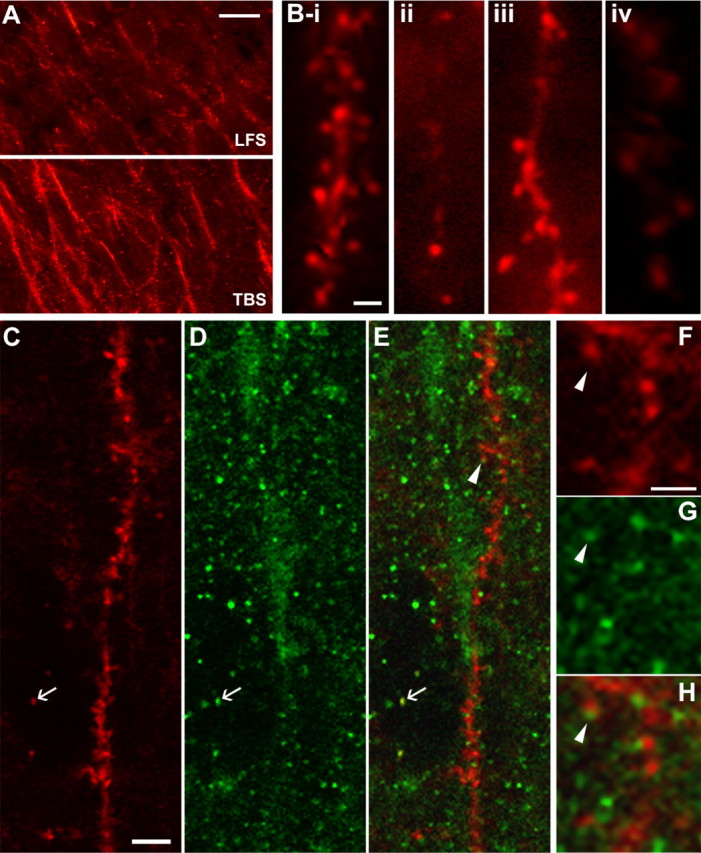
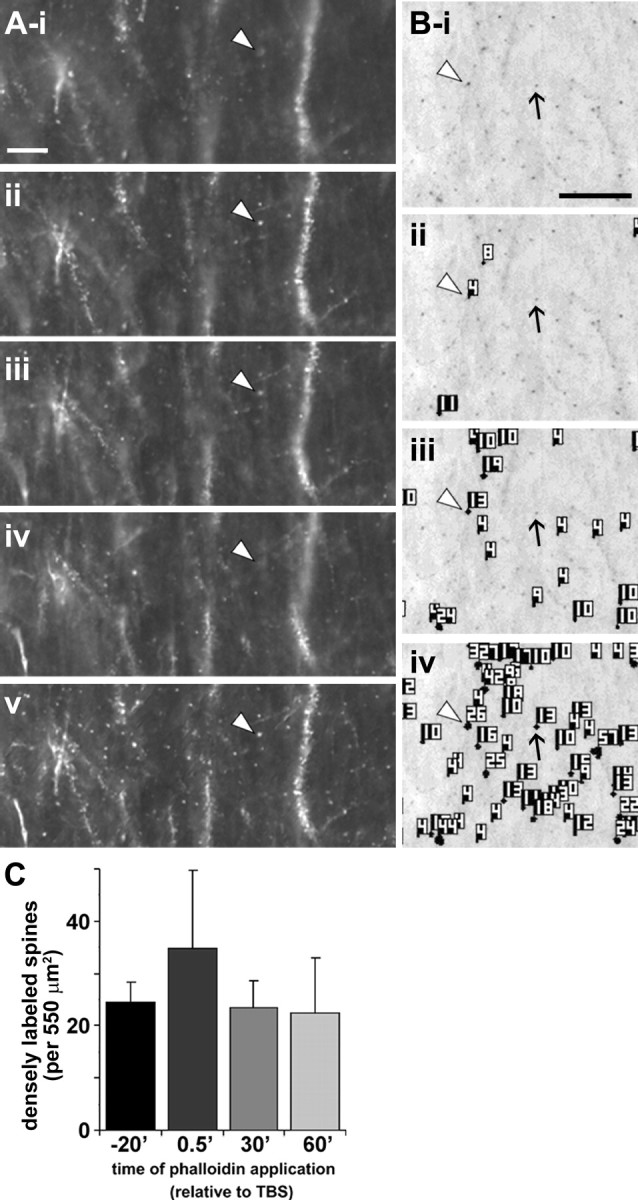
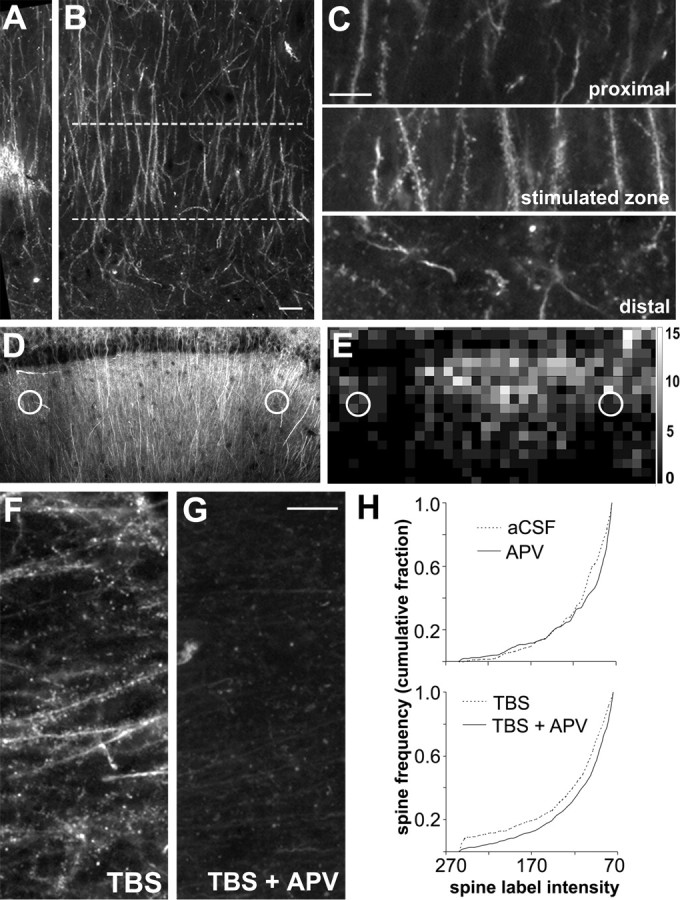
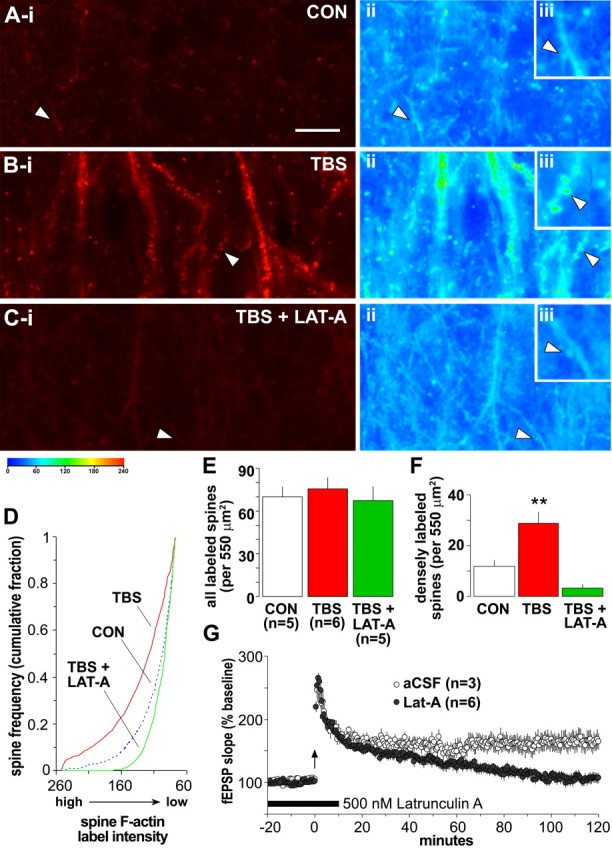
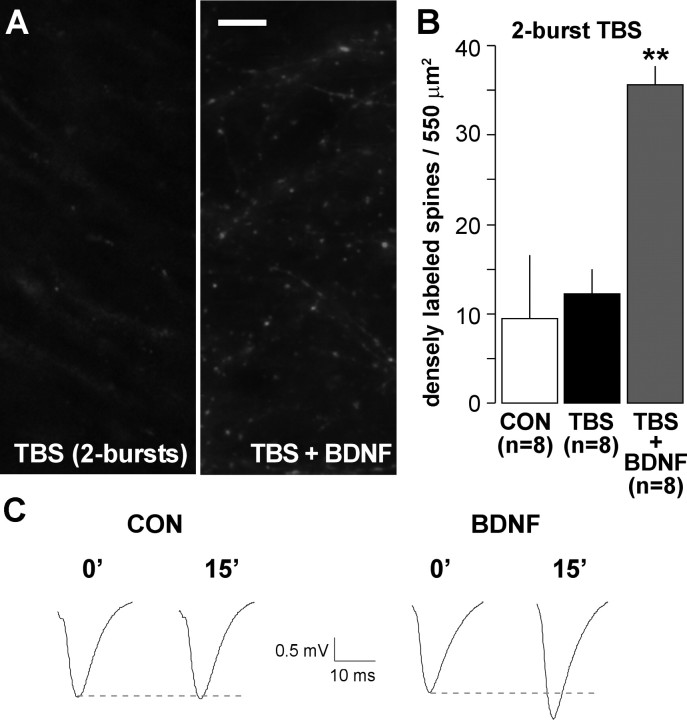
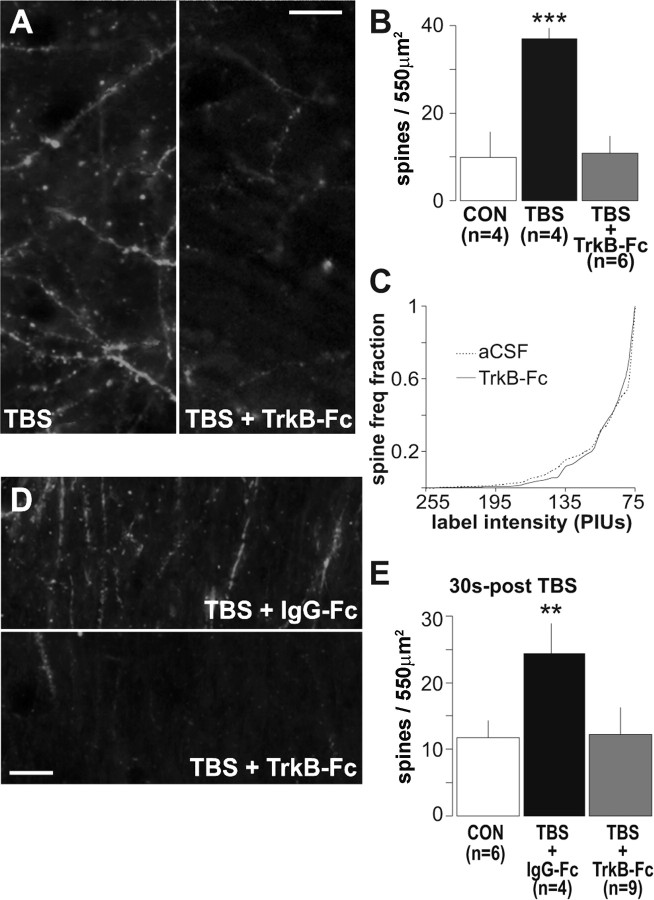
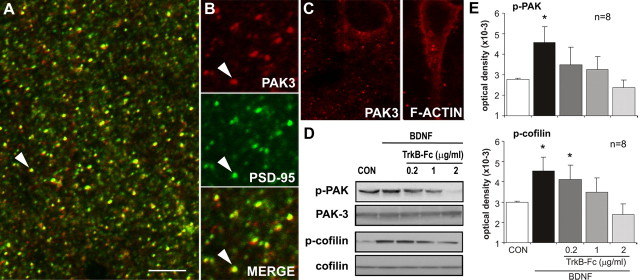
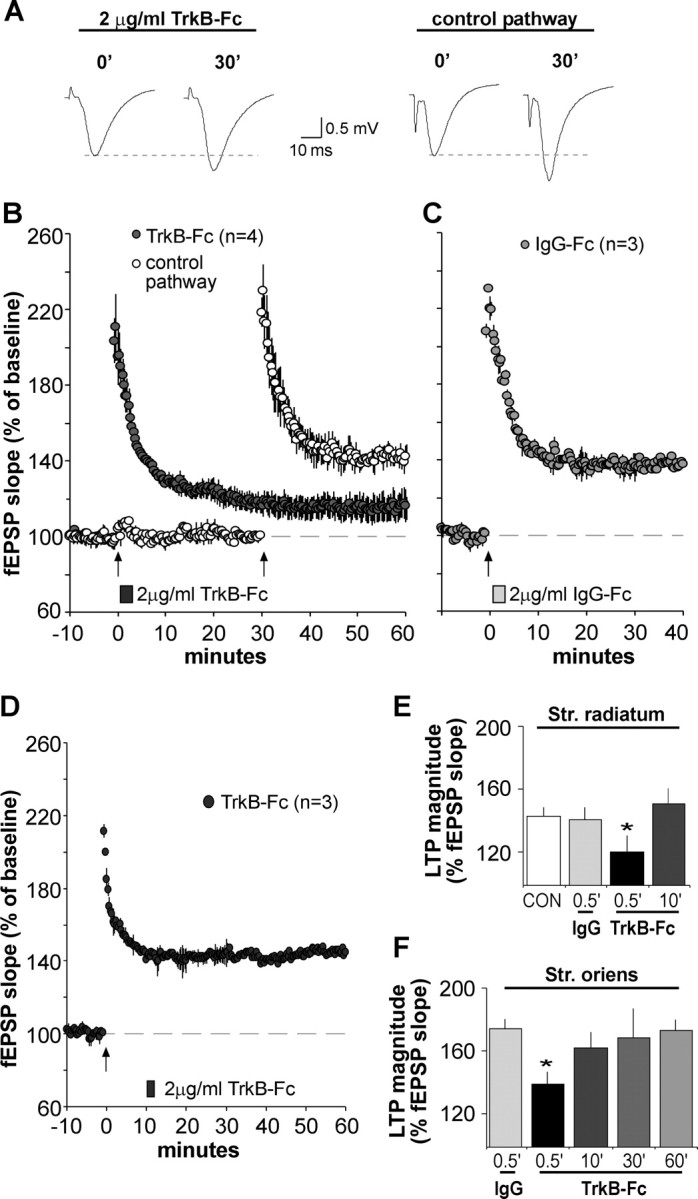
Brain-derived neurotrophic factor (BDNF) is an extremely potent, positive modulator of theta burst induced long-term potentiation (LTP) in the adult hippocampus. The present studies tested whether the neurotrophin exerts its effects by facilitating cytoskeletal changes in dendritic spines. BDNF caused no changes in phalloidin labeling of filamentous actin (F-actin) when applied alone to rat hippocampal slices but markedly enhanced the number of densely labeled spines produced by a threshold level of theta burst stimulation. Conversely, the BDNF scavenger TrkB-Fc completely blocked increases in spine F-actin produced by suprathreshold levels of theta stimulation. TrkB-Fc also blocked LTP consolidation when applied 1-2 min, but not 10 min, after theta trains. Additional experiments confirmed that p21 activated kinase and cofilin, two actin-regulatory proteins implicated in spine morphogenesis, are concentrated in spines in mature hippocampus and further showed that both undergo rapid, dose-dependent phosphorylation after infusion of BDNF. These results demonstrate that the influence of BDNF on the actin cytoskeleton is retained into adulthood in which it serves to positively modulate the time-dependent LTP consolidation process.
Figures
References
-
- Ackermann M, Matus A. Activity-induced targeting of profilin and stabilization of dendritic spine morphology. Nat Neurosci. 2003;6:1194–1200. - PubMed
-
- Allen KM, Gleeson JG, Bagrodia S, Partington MW, MacMillan JC, Cerione RA, Mulley JC, Walsh CA. PAK3 mutation in nonsyndromic X-linked mental retardation. Nat Genet. 1998;20:25–30. - PubMed
-
- Arai A, Larson J, Lynch G. Anoxia reveals a vulnerable period in the development of long-term potentiation. Brain Res. 1990;511:353–357. - PubMed
-
- Arai A, Black J, Lynch G. Origins of the variations in long-term potentiation between synapses in the basal versus apical dendrites of hippocampal neurons. Hippocampus. 1994;4:1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources