

Artificial lymph nodes induce potent secondary immune responses in naive and immunodeficient mice
- PMID: 17364025
- PMCID: PMC1810575
- DOI: 10.1172/JCI30379
Artificial lymph nodes induce potent secondary immune responses in naive and immunodeficient mice
Abstract
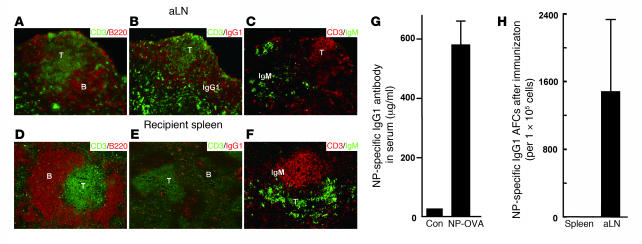
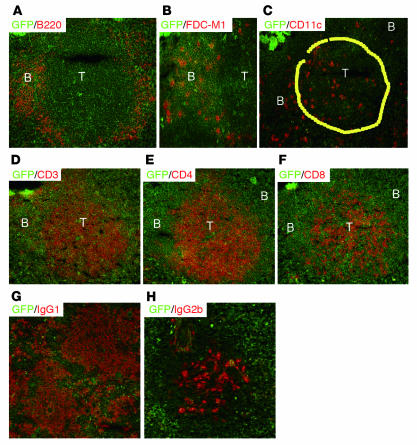
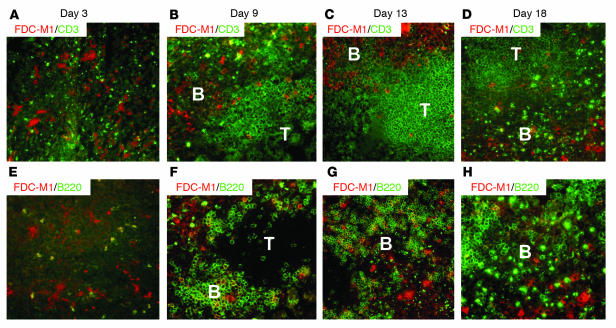
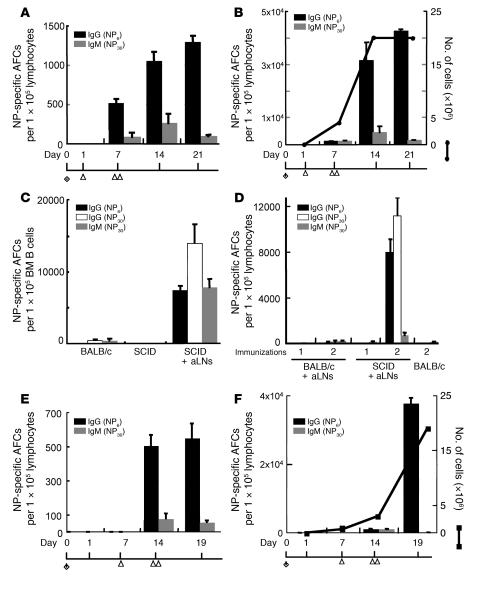
We previously demonstrated that artificial lymph nodes (aLNs) could be generated in mice by the implantation of stromal cell-embedded biocompatible scaffolds into their renal subcapsular spaces. T and B cell domains that form in aLNs have immune response functions similar to those of follicles of normal lymphoid tissue. In the present study, we show that the aLNs were transplantable to normal as well as SCID mice, where they efficiently induced secondary immune responses. Antigen-specific secondary responses were strongly induced in aLNs even 4 weeks after their transplantation. The antigen-specific antibody responses in lymphocyte-deficient SCID mice receiving transplanted aLNs were substantial. The cells from the aLNs migrated to the SCID mouse spleen and BM, where they expanded to generate large numbers of antigen-specific antibody-forming cells. Secondary responses were maintained over time after immunization (i.e., antigen challenge), indicating that aLNs can support the development of memory B cells and long-lived plasma cells. Memory CD4(+) T cells were enriched in the aLNs and spleens of aLN-transplanted SCID mice. Our results indicate that aLNs support strong antigen-specific secondary antibody responses in immunodeficient mice and suggest the possibility of future clinical applications.
Figures
Similar articles
-
[Generation of artificial lymph nodes (aLNs) and their immunological function].Nihon Rinsho. 2008 Jul;66(7):1391-6. Nihon Rinsho. 2008. PMID: 18616133 Review. Japanese.
-
Generation of a synthetic lymphoid tissue-like organoid in mice.Nat Biotechnol. 2004 Dec;22(12):1539-45. doi: 10.1038/nbt1039. Epub 2004 Nov 28. Nat Biotechnol. 2004. PMID: 15568019
-
Immunological memory function of the T and B cell types: distribution over mouse spleen and lymph nodes.Eur J Immunol. 1975 Mar;5(3):211-5. doi: 10.1002/eji.1830050312. Eur J Immunol. 1975. PMID: 1086225
-
Lymphoid bone marrow cultures can reconstitute heterogeneous B and T cell-dependent responses in severe combined immunodeficient mice.J Immunol. 1986 Dec 1;137(11):3457-63. J Immunol. 1986. PMID: 2878043
-
Popliteal lymph node assay: facts and perspectives.J Appl Toxicol. 2005 Nov-Dec;25(6):451-8. doi: 10.1002/jat.1072. J Appl Toxicol. 2005. PMID: 15986413 Review.
Cited by
-
Bioengineering of Artificial Lymphoid Organs.Acta Naturae. 2016 Apr-Jun;8(2):10-23. Acta Naturae. 2016. PMID: 27437136 Free PMC article.
-
Selective interaction of PEGylated polyglutamic acid nanocapsules with cancer cells in a 3D model of a metastatic lymph node.J Nanobiotechnology. 2016 Jun 23;14(1):51. doi: 10.1186/s12951-016-0207-8. J Nanobiotechnology. 2016. PMID: 27339609 Free PMC article.
-
Lymphatic Tissue and Organ Engineering for In Vitro Modeling and In Vivo Regeneration.Cold Spring Harb Perspect Med. 2022 Mar 14;12(9):a041169. doi: 10.1101/cshperspect.a041169. Online ahead of print. Cold Spring Harb Perspect Med. 2022. PMID: 35288402 Free PMC article.
-
Gel-Trapped Lymphorganogenic Chemokines Trigger Artificial Tertiary Lymphoid Organs and Mount Adaptive Immune Responses In Vivo.Front Immunol. 2016 Aug 22;7:316. doi: 10.3389/fimmu.2016.00316. eCollection 2016. Front Immunol. 2016. PMID: 27597851 Free PMC article.
-
Tertiary lymphoid structural heterogeneity determines tumour immunity and prospects for clinical application.Mol Cancer. 2024 Apr 6;23(1):75. doi: 10.1186/s12943-024-01980-6. Mol Cancer. 2024. PMID: 38582847 Free PMC article. Review.
References
-
- von Andrian U.H., Mempel T.R. Homing and cellular traffic in lymph nodes. Nat. Rev. Immunol. 2003;3:867–878. - PubMed
-
- Miyasaka M., Tanaka T. Lymphocyte trafficking across high endothelial venules: dogmas and enigmas. Nat. Rev. Immunol. 2004;4:360–370. - PubMed
-
- De Togni P., et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science. 1994;264:703–707. - PubMed
-
- Banks T.A., et al. Lymphotoxin-alpha-deficient mice. Effects on secondary lymphoid organ development and humoral immune responsiveness. J. Immunol. . 1995;155:1685–1693. - PubMed
-
- Fu Y.X., Chaplin D.D. Development and maturation of secondary lymphoid tissues. Annu. Rev. Immunol. 1999;17:399–433. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
