

A conformational switch in the ligand-binding domain regulates the dependence of the glucocorticoid receptor on Hsp90
- PMID: 17367809
- PMCID: PMC2596751
- DOI: 10.1016/j.jmb.2007.02.057
A conformational switch in the ligand-binding domain regulates the dependence of the glucocorticoid receptor on Hsp90
Abstract
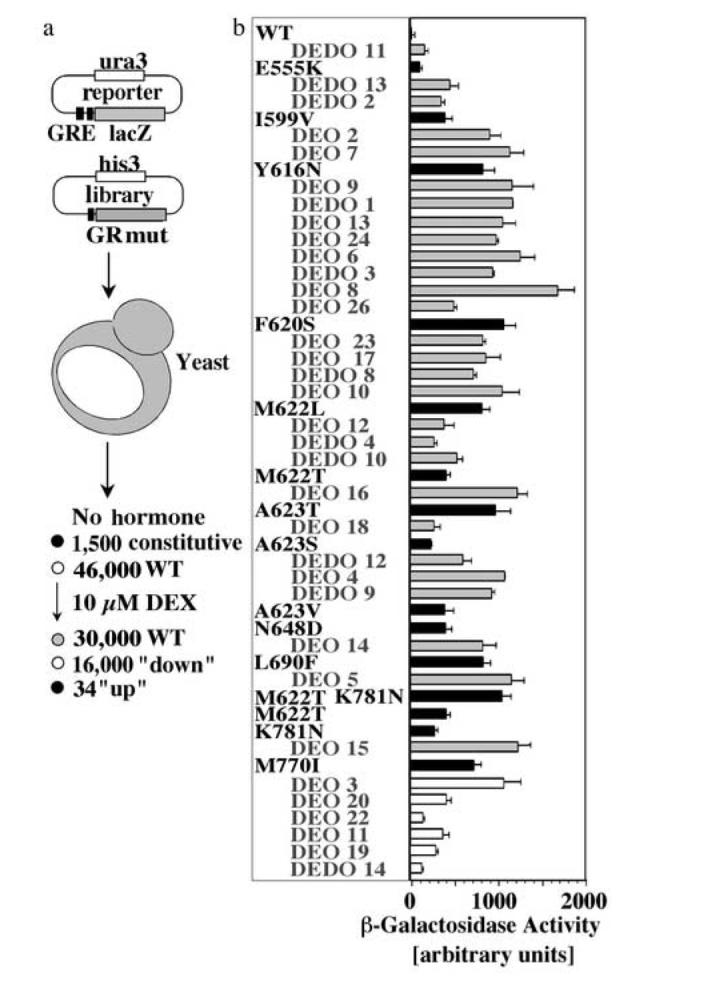
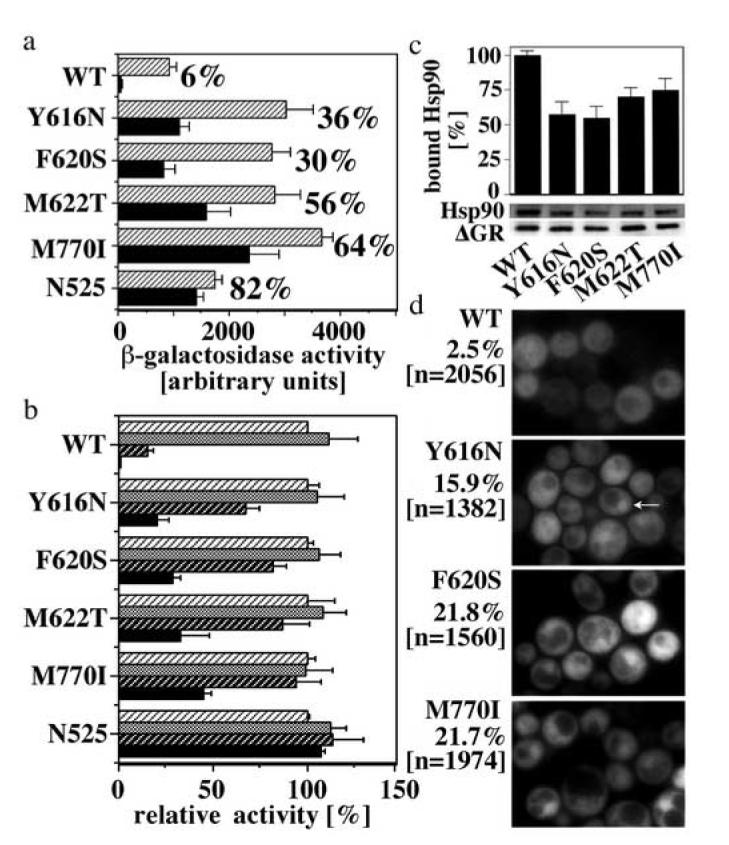
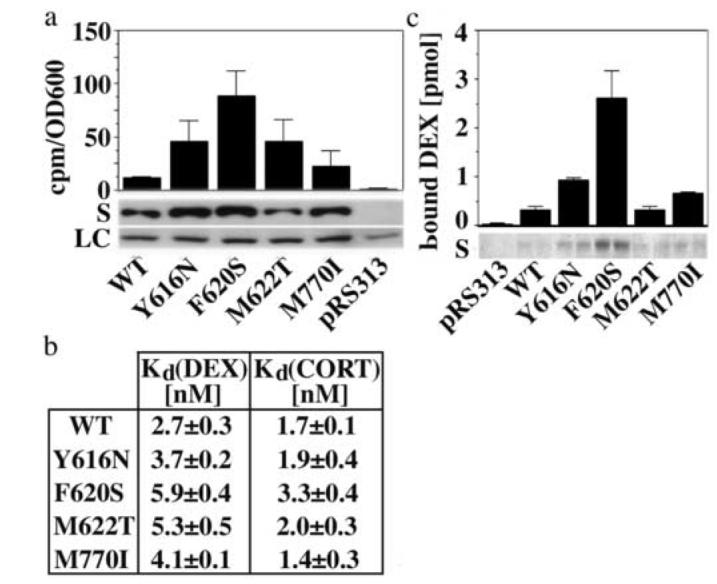
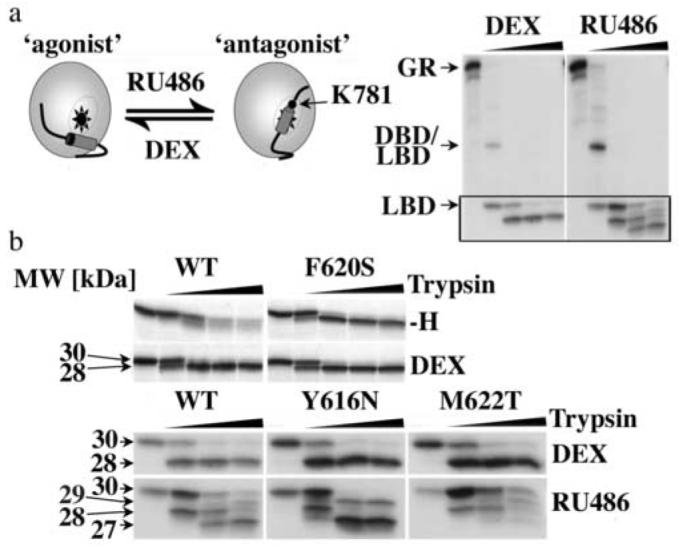
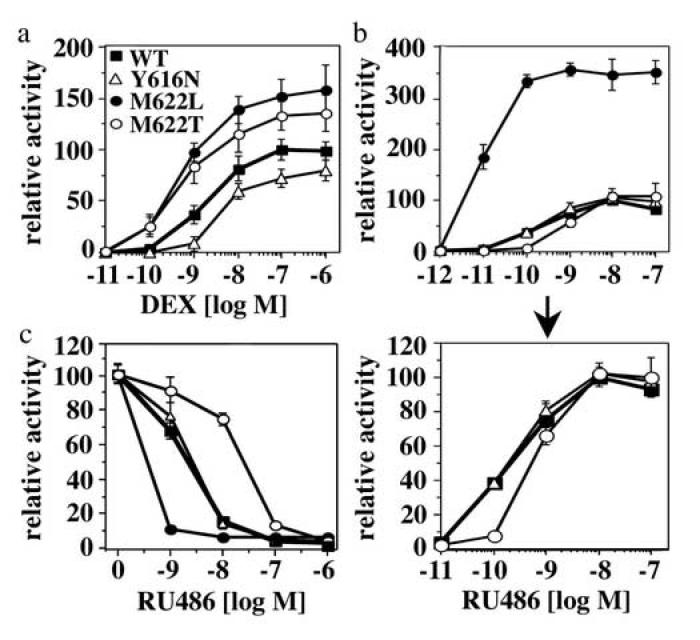
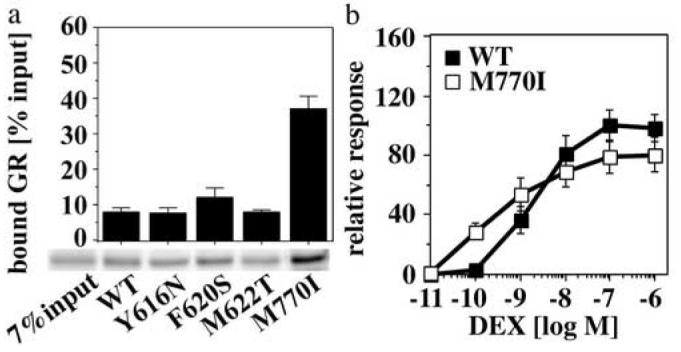
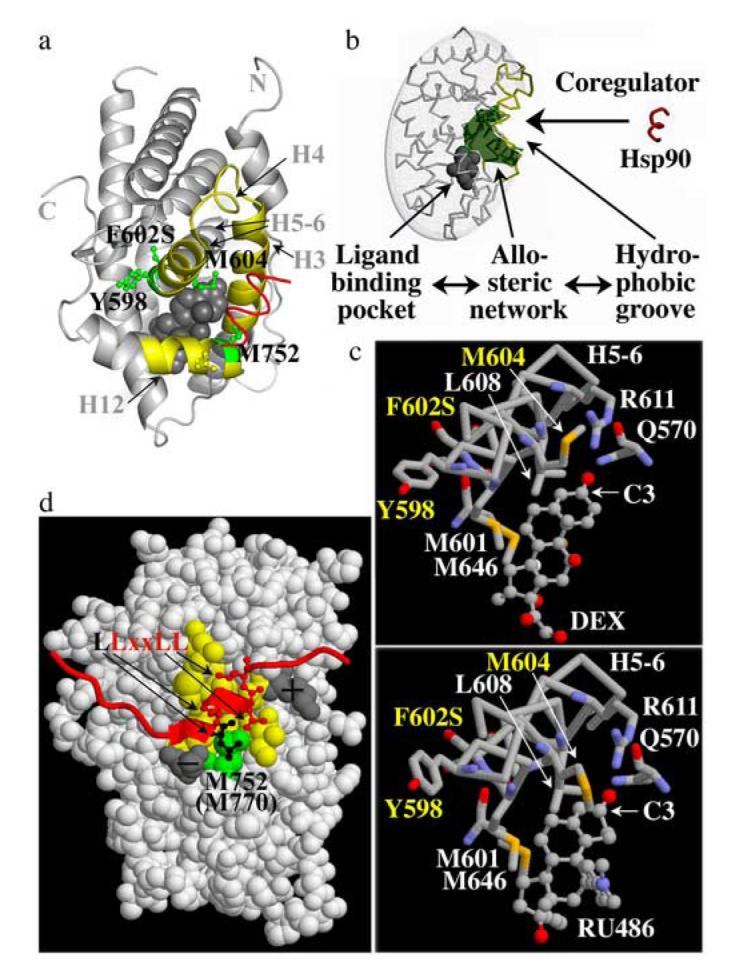
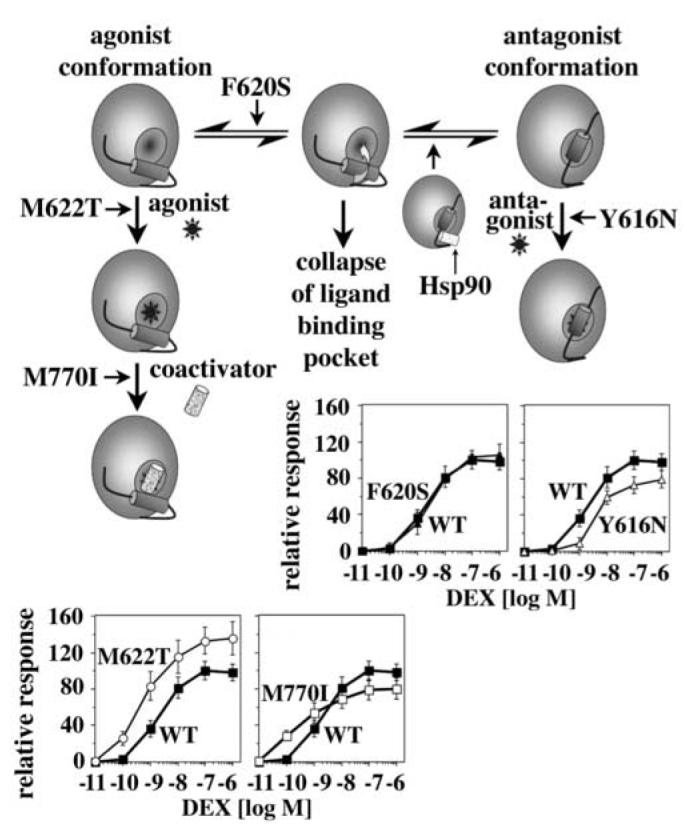
Steroid hormone receptors (SRs) are transcription factors that act as regulatory switches by altering gene expression in response to ligands. The highly conserved ligand-binding domain of SRs is a precise but versatile molecular switch that can adopt distinct conformations. Differential stabilization of these conformations by ligands, DNA response elements and transcriptional coregulators controls the activity of SRs in a gene-specific and cell-specific manner. In the case of the glucocorticoid receptor (GR), high-affinity ligand binding requires the interaction of the LBD with the heat shock protein 90 (Hsp90). Here, we show that the dependence of the ligand binding ability of GR on Hsp90 can be modified by the replacement of single amino acids within an allosteric network that connects the buried ligand-binding pocket and a solvent-exposed coregulator interaction surface. Each of the identified mutations altered the equilibrium between alternative GR conformations distinctively, indicating that the Hsp90 dependence of SRs may correlate with differences in the conformational dynamics of these receptors. Our results suggest that Hsp90 stabilizes the GR ligand-binding pocket indirectly by utilizing the allosteric network, while allowing the receptor to remain structurally uncommitted. Thus, in addition to ensuring the accessibility of the GR ligand-binding pocket to ligands, Hsp90 seems to enable hormones and coregulators to act as allosteric effectors, which forms the basis for gene-specific and cell-specific responses of GR to ligands.
Figures
References
-
- Aranda A, Pascual A. Nuclear hormone receptors and gene expression. Physiol Rev. 2001;81:1269–304. - PubMed
-
- Robin-Jagerschmidt C, Wurtz JM, Guillot B, Gofflo D, Benhamou B, Vergezac A, Ossart C, Moras D, Philibert D. Residues in the ligand binding domain that confer progestin or glucocorticoid specificity and modulate the receptor transactivation capacity. Mol Endocrinol. 2000;14:1028–37. - PubMed
-
- Nettles KW, Sun J, Radek JT, Sheng S, Rodriguez AL, Katzenellenbogen JA, Katzenellenbogen BS, Greene GL. Allosteric control of ligand selectivity between estrogen receptors alpha and beta: implications for other nuclear receptors. Mol Cell. 2004;13:317–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
