

Influence of norepinephrine on somatosensory neuronal responses in the rat thalamus: a combined modeling and in vivo multi-channel, multi-neuron recording study
- PMID: 17368434
- PMCID: PMC4529675
- DOI: 10.1016/j.brainres.2007.02.006
Influence of norepinephrine on somatosensory neuronal responses in the rat thalamus: a combined modeling and in vivo multi-channel, multi-neuron recording study
Abstract
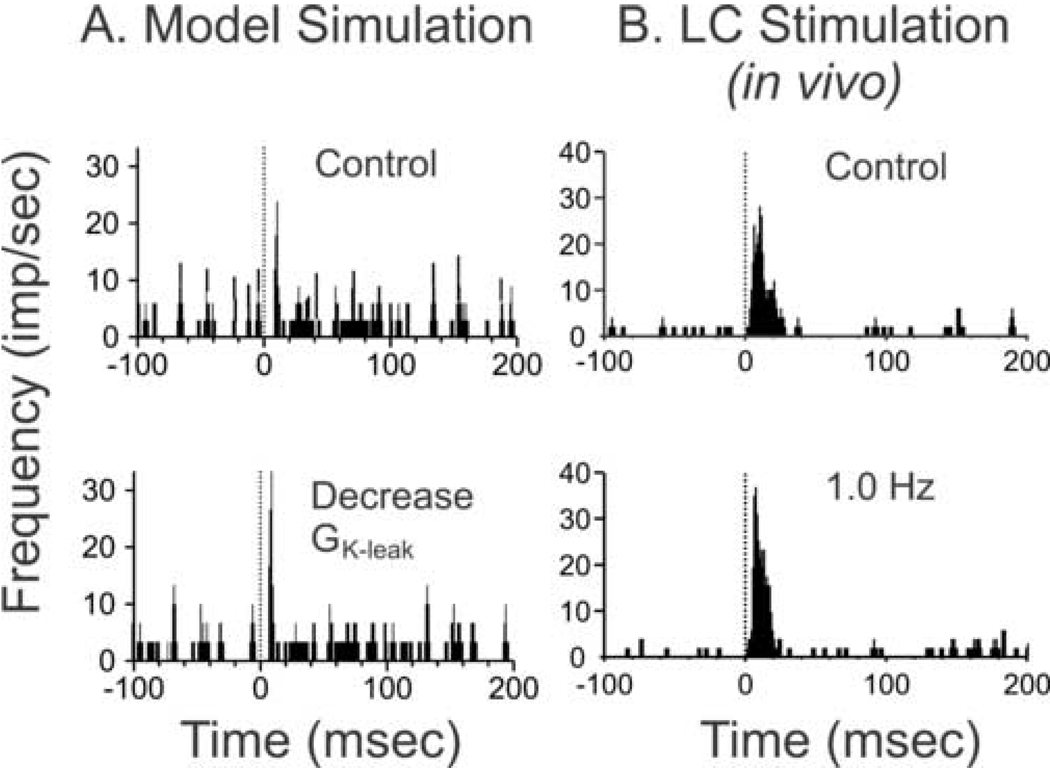
Norepinephrine released within primary sensory circuits from locus coeruleus afferent fibers can produce a spectrum of modulatory actions on spontaneous or sensory-evoked activity of individual neurons. Within the ventral posterior medial thalamus, membrane currents modulated by norepinephrine have been identified. However, the relationship between the cellular effects of norepinephrine and the impact of norepinephrine release on populations of neurons encoding sensory signals is still open to question. To address this lacuna in understanding the net impact of the noradrenergic system on sensory signal processing, a computational model of the rat trigeminal somatosensory thalamus was generated. The effects of independent manipulation of different cellular actions of norepinephrine on simulated afferent input to the computational model were then examined. The results of these simulations aided in the design of in vivo neural ensemble recording experiments where sensory-driven responses of thalamic neurons were measured before and during locus coeruleus activation in waking animals. Together the simulated and experimental results reveal several key insights regarding the regulation of neural network operation by norepinephrine including: 1) cell-specific modulatory actions of norepinephrine, 2) mechanisms of norepinephrine action that can improve the tuning of the network and increase the signal-to-noise ratio of cellular responses in order to enhance network representation of salient stimulus features and 3) identification of the dynamic range of thalamic neuron function through which norepinephrine operates.
Figures





References
-
- Akaike T. Periodic bursting activities of locus coerulleus neurons in the rat. Brain Res. 1982;239:629–633. - PubMed
-
- Bal T, McCormick DA. What stops synchronized thalamocortical oscillations? Neuron. 1996;17:297–308. - PubMed
-
- Bazhenov M, Timofeev I, Steriade M, Sejnowski T. Spiking-bursting activity in the thalamic reticular nucleus initiates sequences of spindle oscillations in thalamic networks. J Neurophysiol. 2000;84:1076–1087. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
