

The SsrA-SmpB ribosome rescue system is important for growth of Bacillus subtilis at low and high temperatures
- PMID: 17369301
- PMCID: PMC1913333
- DOI: 10.1128/JB.00062-07
The SsrA-SmpB ribosome rescue system is important for growth of Bacillus subtilis at low and high temperatures
Abstract
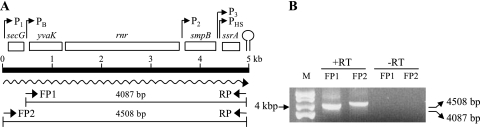
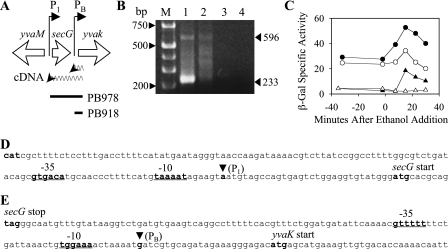
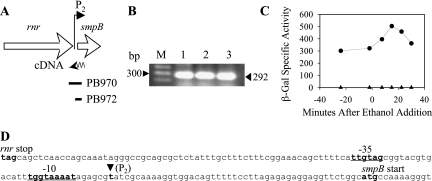
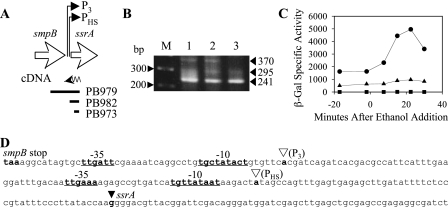
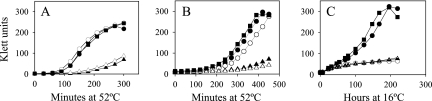
Bacillus subtilis has multiple stress response systems whose integrated action promotes growth and survival under unfavorable conditions. Here we address the function and transcriptional organization of a five-gene cluster containing ssrA, previously known to be important for growth at high temperature because of the role of its tmRNA product in rescuing stalled ribosomes. Reverse transcription-PCR experiments detected a single message for the secG-yvaK-rnr-smpB-ssrA cluster, suggesting that it constitutes an operon. However, rapid amplification of cDNA ends-PCR and lacZ fusion experiments indicated that operon transcription is complex, with at least five promoters controlling different segments of the cluster. One sigma(A)-like promoter preceded secG (P(1)), and internal sigma(A)-like promoters were found in both the rnr-smpB (P(2)) and smpB-ssrA intervals (P(3) and P(HS)). Another internal promoter lay in the secG-yvaK intercistronic region, and this activity (P(B)) was dependent on the general stress factor sigma(B). Null mutations in the four genes downstream from P(B) were tested for their effects on growth. Loss of yvaK (carboxylesterase E) or rnr (RNase R) caused no obvious phenotype. By contrast, smpB was required for growth at high temperature (52 degrees C), as anticipated if its product (a small ribosomal binding protein) is essential for tmRNA (ssrA) function. Notably, smpB and ssrA were also required for growth at low temperature (16 degrees C), a phenotype not previously associated with tmRNA activity. These results extend the known high-temperature role of ssrA and indicate that the ribosome rescue system is important at both extremes of the B. subtilis temperature range.
Figures
Similar articles
-
Requirement of transfer-messenger RNA for the growth of Bacillus subtilis under stresses.Genes Cells. 2000 Aug;5(8):627-35. doi: 10.1046/j.1365-2443.2000.00356.x. Genes Cells. 2000. PMID: 10947848
-
Four additional genes in the sigB operon of Bacillus subtilis that control activity of the general stress factor sigma B in response to environmental signals.J Bacteriol. 1995 Jan;177(1):123-33. doi: 10.1128/jb.177.1.123-133.1995. J Bacteriol. 1995. PMID: 8002610 Free PMC article.
-
Mycobacterium tuberculosis possesses an unusual tmRNA rescue system.Tuberculosis (Edinb). 2014 Jan;94(1):34-42. doi: 10.1016/j.tube.2013.09.007. Epub 2013 Oct 9. Tuberculosis (Edinb). 2014. PMID: 24145139
-
The SsrA-SmpB system for protein tagging, directed degradation and ribosome rescue.Nat Struct Biol. 2000 Jun;7(6):449-55. doi: 10.1038/75843. Nat Struct Biol. 2000. PMID: 10881189 Review.
-
An overview of the bacterial SsrA system modulating intracellular protein levels and activities.Appl Microbiol Biotechnol. 2020 Jun;104(12):5229-5241. doi: 10.1007/s00253-020-10623-x. Epub 2020 Apr 28. Appl Microbiol Biotechnol. 2020. PMID: 32342145 Review.
Cited by
-
Mechanisms of ribosome rescue in bacteria.Nat Rev Microbiol. 2015 May;13(5):285-97. doi: 10.1038/nrmicro3438. Epub 2015 Apr 13. Nat Rev Microbiol. 2015. PMID: 25874843 Review.
-
Comparative genomic analysis of seven Mycoplasma hyosynoviae strains.Microbiologyopen. 2015 Apr;4(2):343-359. doi: 10.1002/mbo3.242. Epub 2015 Feb 18. Microbiologyopen. 2015. PMID: 25693846 Free PMC article.
-
Biochemical and Cellular Characterization of the Function of Fluorophosphonate-Binding Hydrolase H (FphH) in Staphylococcus aureus Support a Role in Bacterial Stress Response.ACS Infect Dis. 2023 Nov 10;9(11):2119-2132. doi: 10.1021/acsinfecdis.3c00246. Epub 2023 Oct 12. ACS Infect Dis. 2023. PMID: 37824340 Free PMC article.
-
Similar but Distinct-Biochemical Characterization of the Staphylococcus aureus Serine Hydrolases FphH and FphI.Proteins. 2025 May;93(5):1009-1021. doi: 10.1002/prot.26785. Epub 2024 Dec 26. Proteins. 2025. PMID: 39726198
-
Francisella tularensis tmRNA system mutants are vulnerable to stress, avirulent in mice, and provide effective immune protection.Mol Microbiol. 2012 Jul;85(1):122-41. doi: 10.1111/j.1365-2958.2012.08093.x. Epub 2012 May 28. Mol Microbiol. 2012. PMID: 22571636 Free PMC article.
References
-
- Band, L., H. Shimotsu, and D. J. Henner. 1984. Nucleotide sequence of the Bacillus subtilis trpE and trpD genes. Gene 27:55-65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
