

On the mechanism of TBA block of the TRPV1 channel
- PMID: 17369424
- PMCID: PMC1868982
- DOI: 10.1529/biophysj.106.102400
On the mechanism of TBA block of the TRPV1 channel
Abstract
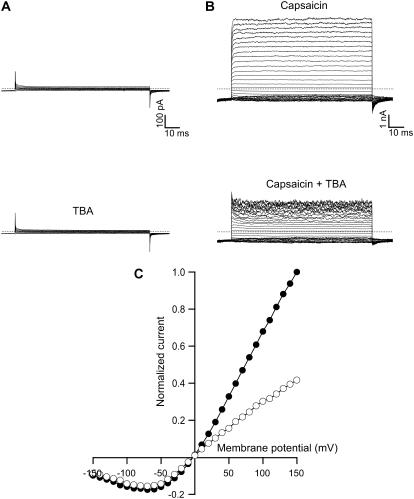
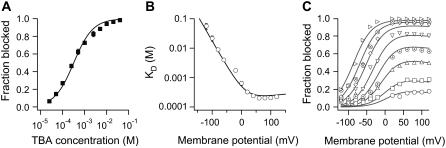
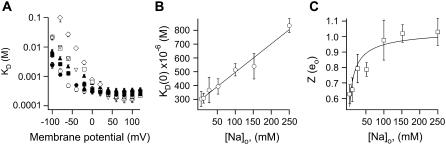
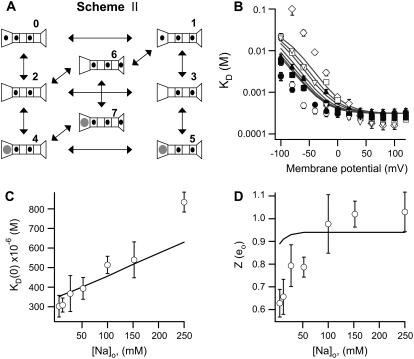
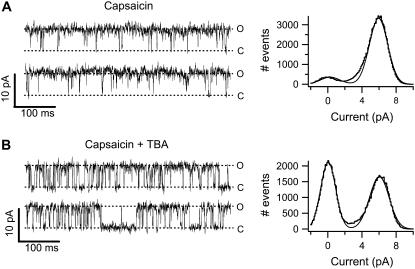
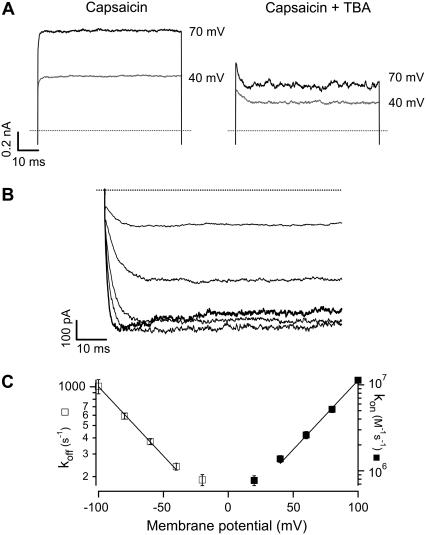
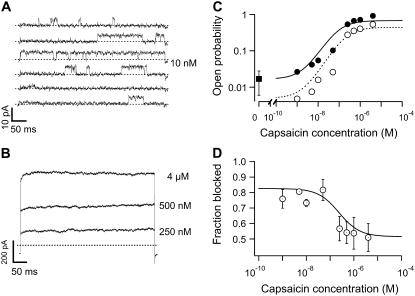
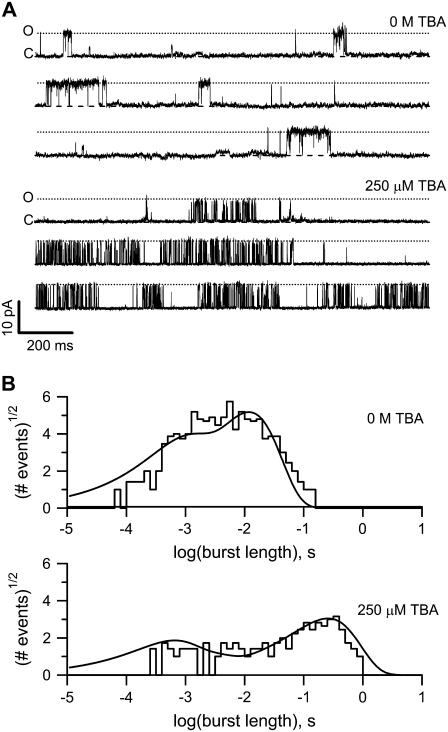
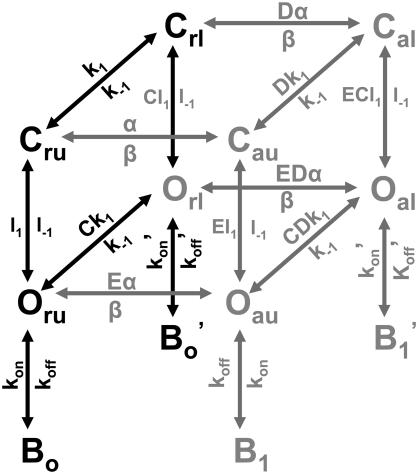
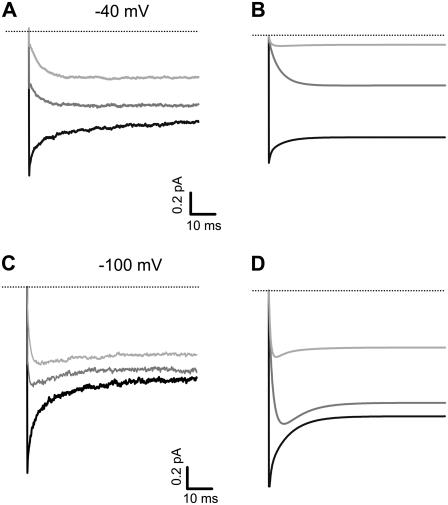
The transient receptor potential vanilloid 1 (TRPV1) channel is a nonselective cation channel activated by capsaicin and responsible for thermosensation. To date, little is known about the gating characteristics of these channels. Here we used tetrabutylammonium (TBA) to determine whether this molecule behaves as an ion conduction blocker in TRPV1 channels and to gain insight into the nature of the activation gate of this protein. TBA belongs to a family of classic potassium channel blockers that have been widely used as tools for determining the localization of the activation gate and the properties of the pore of several ion channels. We found TBA to be a voltage-dependent pore blocker and that the properties of block are consistent with an open-state blocker, with the TBA molecule binding to multiple open states, each with different blocker affinities. Kinetics of channel closure and burst-length analysis in the presence of blocker are consistent with a state-dependent blocking mechanism, with TBA interfering with closing of an activation gate. This activation gate may be located cytoplasmically with respect to the binding site of TBA ions, similar to what has been observed in potassium channels. We propose an allosteric model for TRPV1 activation and block by TBA, which explains our experimental data.
Figures
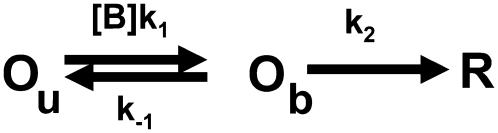
Similar articles
-
Properties of the inner pore region of TRPV1 channels revealed by block with quaternary ammoniums.J Gen Physiol. 2008 Nov;132(5):547-62. doi: 10.1085/jgp.200810051. J Gen Physiol. 2008. PMID: 18955595 Free PMC article.
-
New insights on the voltage dependence of the KCa3.1 channel block by internal TBA.J Gen Physiol. 2004 Oct;124(4):333-48. doi: 10.1085/jgp.200409145. J Gen Physiol. 2004. PMID: 15452196 Free PMC article.
-
Mechanism of intracellular block of the KcsA K+ channel by tetrabutylammonium: insights from X-ray crystallography, electrophysiology and replica-exchange molecular dynamics simulations.J Mol Biol. 2007 Jan 19;365(3):649-62. doi: 10.1016/j.jmb.2006.09.069. Epub 2006 Sep 29. J Mol Biol. 2007. PMID: 17070844
-
ThermoTRP channels as modular proteins with allosteric gating.Cell Calcium. 2007 Oct-Nov;42(4-5):427-38. doi: 10.1016/j.ceca.2007.04.004. Epub 2007 May 17. Cell Calcium. 2007. PMID: 17499848 Review.
-
Capsaicin receptor: TRPV1 a promiscuous TRP channel.Handb Exp Pharmacol. 2007;(179):155-71. doi: 10.1007/978-3-540-34891-7_9. Handb Exp Pharmacol. 2007. PMID: 17217056 Review.
Cited by
-
Structural mechanisms of transient receptor potential ion channels.J Gen Physiol. 2020 Mar 2;152(3):e201811998. doi: 10.1085/jgp.201811998. J Gen Physiol. 2020. PMID: 31972006 Free PMC article. Review.
-
An external sodium ion binding site controls allosteric gating in TRPV1 channels.Elife. 2016 Feb 12;5:e13356. doi: 10.7554/eLife.13356. Elife. 2016. PMID: 26882503 Free PMC article.
-
Nobilamides A-H, long-acting transient receptor potential vanilloid-1 (TRPV1) antagonists from mollusk-associated bacteria.J Med Chem. 2011 Jun 9;54(11):3746-55. doi: 10.1021/jm101621u. Epub 2011 May 16. J Med Chem. 2011. PMID: 21524089 Free PMC article.
-
NGF-mediated crosstalk: unraveling the influence of metabolic deregulation on the interplay between neural and pancreatic cancer cells and its impact on patient outcomes.Front Pharmacol. 2024 Dec 11;15:1499414. doi: 10.3389/fphar.2024.1499414. eCollection 2024. Front Pharmacol. 2024. PMID: 39723256 Free PMC article. Review.
-
N-oleoyldopamine, a novel endogenous capsaicin-like lipid, protects the heart against ischemia-reperfusion injury via activation of TRPV1.Am J Physiol Heart Circ Physiol. 2008 Aug;295(2):H728-35. doi: 10.1152/ajpheart.00022.2008. Epub 2008 Jun 20. Am J Physiol Heart Circ Physiol. 2008. PMID: 18567714 Free PMC article.
References
-
- Minke, B. 1977. Drosophila mutant with a transducer defect. Biophys. Struct. Mech. 3:59–64. - PubMed
-
- Clapham, D. E., L. W. Runnels, and C. Strubing. 2001. The TRP ion channel family. Nat. Rev. Neurosci. 2:387–396. - PubMed
-
- Montell, C. 2001. Physiology, phylogeny, and functions of the TRP superfamily of cation channels. Sci. STKE. 2001:RE1. - PubMed
-
- Montell, C., L. Birnbaumer, V. Flockerzi, R. J. Bindels, E. A. Bruford, M. J. Caterina, D. E. Clapham, C. Harteneck, S. Heller, D. Julius, I. Kojima, Y. Mori, R. Penner, D. Prawitt, A. M. Scharenberg, G. Schultz, N. Shimizu, and M. X. Zhu. 2002. A unified nomenclature for the superfamily of TRP cation channels. Mol. Cell. 9:229–231. - PubMed
-
- Caterina, M. J., M. A. Schumacher, M. Tominaga, T. A. Rosen, J. D. Levine, and D. Julius. 1997. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 389:816–824. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
