

MAM3 catalyzes the formation of all aliphatic glucosinolate chain lengths in Arabidopsis
- PMID: 17369439
- PMCID: PMC1913788
- DOI: 10.1104/pp.106.091579
MAM3 catalyzes the formation of all aliphatic glucosinolate chain lengths in Arabidopsis
Abstract
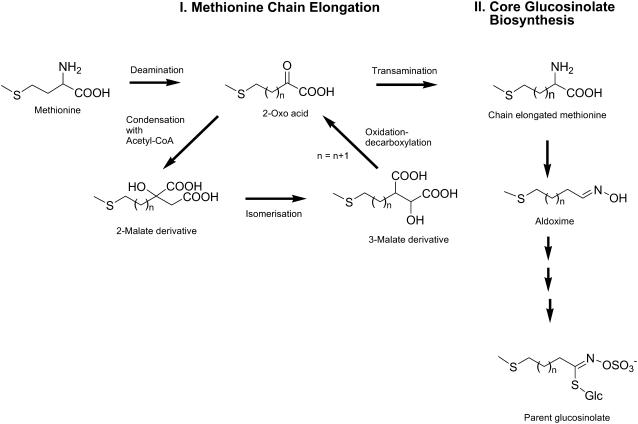
Chain elongated, methionine (Met)-derived glucosinolates are a major class of secondary metabolites in Arabidopsis (Arabidopsis thaliana). The key enzymatic step in determining the length of the chain is the condensation of acetyl-coenzyme A with a series of omega-methylthio-2-oxoalkanoic acids, catalyzed by methylthioalkylmalate (MAM) synthases. The existence of two MAM synthases has been previously reported in the Arabidopsis ecotype Columbia: MAM1 and MAM3 (formerly known as MAM-L). Here, we describe the biochemical properties of the MAM3 enzyme, which is able to catalyze all six condensation reactions of Met chain elongation that occur in Arabidopsis. Underlining its broad substrate specificity, MAM3 also accepts a range of non-Met-derived 2-oxoacids, e.g. converting pyruvate to citramalate and 2-oxoisovalerate to isopropylmalate, a step in leucine biosynthesis. To investigate its role in vivo, we identified plant lines with mutations in MAM3 that resulted in a complete lack or greatly reduced levels of long-chain glucosinolates. This phenotype could be complemented by reintroduction of a MAM3 expression construct. Analysis of MAM3 mutants demonstrated that MAM3 catalyzes the formation of all glucosinolate chain lengths in vivo as well as in vitro, making this enzyme the major generator of glucosinolate chain length diversity in the plant. The localization of MAM3 in the chloroplast suggests that this organelle is the site of Met chain elongation.
Figures




Similar articles
-
Expression pattern of the glucosinolate side chain biosynthetic genes MAM1 and MAM3 of Arabidopsis thaliana in different organs and developmental stages.Plant Physiol Biochem. 2012 Apr;53:77-83. doi: 10.1016/j.plaphy.2012.01.015. Epub 2012 Jan 28. Plant Physiol Biochem. 2012. PMID: 22336876
-
Changing substrate specificity and iteration of amino acid chain elongation in glucosinolate biosynthesis through targeted mutagenesis of Arabidopsis methylthioalkylmalate synthase 1.Biosci Rep. 2019 Jul 2;39(7):BSR20190446. doi: 10.1042/BSR20190446. Print 2019 Jul 31. Biosci Rep. 2019. PMID: 31175145 Free PMC article.
-
Arabidopsis thaliana encodes a bacterial-type heterodimeric isopropylmalate isomerase involved in both Leu biosynthesis and the Met chain elongation pathway of glucosinolate formation.Plant Mol Biol. 2009 Oct;71(3):227-39. doi: 10.1007/s11103-009-9519-5. Epub 2009 Jul 14. Plant Mol Biol. 2009. PMID: 19597944 Free PMC article.
-
Biosynthesis of methionine-derived glucosinolates in Arabidopsis thaliana: recombinant expression and characterization of methylthioalkylmalate synthase, the condensing enzyme of the chain-elongation cycle.Planta. 2004 Apr;218(6):1026-35. doi: 10.1007/s00425-003-1184-3. Epub 2004 Jan 22. Planta. 2004. PMID: 14740211
-
Biosynthesis and metabolic engineering of glucosinolates.Amino Acids. 2002;22(3):279-95. doi: 10.1007/s007260200014. Amino Acids. 2002. PMID: 12083070 Review.
Cited by
-
Reduced glucosinolate content in oilseed rape (Brassica napus L.) by random mutagenesis of BnMYB28 and BnCYP79F1 genes.Sci Rep. 2023 Feb 9;13(1):2344. doi: 10.1038/s41598-023-28661-6. Sci Rep. 2023. PMID: 36759657 Free PMC article.
-
A systems biology approach identifies a R2R3 MYB gene subfamily with distinct and overlapping functions in regulation of aliphatic glucosinolates.PLoS One. 2007 Dec 19;2(12):e1322. doi: 10.1371/journal.pone.0001322. PLoS One. 2007. PMID: 18094747 Free PMC article.
-
Effects of Sulfur Assimilation in Pseudomonas fluorescens SS101 on Growth, Defense, and Metabolome of Different Brassicaceae.Biomolecules. 2021 Nov 16;11(11):1704. doi: 10.3390/biom11111704. Biomolecules. 2021. PMID: 34827700 Free PMC article.
-
ESKIMO1 is a key gene involved in water economy as well as cold acclimation and salt tolerance.BMC Plant Biol. 2008 Dec 7;8:125. doi: 10.1186/1471-2229-8-125. BMC Plant Biol. 2008. PMID: 19061521 Free PMC article.
-
Comparative transcriptomic analyses of glucosinolate metabolic genes during the formation of Chinese kale seeds.BMC Plant Biol. 2021 Aug 21;21(1):394. doi: 10.1186/s12870-021-03168-2. BMC Plant Biol. 2021. PMID: 34418959 Free PMC article.
References
-
- Bechtold N, Ellis J, Pelletier G (1993) In-planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci Paris Life Sci 316 1194–1199
-
- Brown PD, Tokuhisa JG, Reichelt M, Gershenzon J (2003) Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 62 471–481 - PubMed
-
- Campos de Quiros H, Magrath R, McCallum D, Kroymann J, Schnabelrauch D, Mitchell-Olds T, Richard M (2000) α-Keto acid elongation and glucosinolate biosynthesis in Arabidopsis thaliana. Theor Appl Genet 101 429–437
-
- Chapple CCS, Decicco C, Ellis BE (1988) Biosynthesis of 2-(2′-methylthio)ethylmalate in Brassica carinata. Phytochemistry 27 3461–3463
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
