

Fibulin-5/DANCE has an elastogenic organizer activity that is abrogated by proteolytic cleavage in vivo
- PMID: 17371835
- PMCID: PMC2064089
- DOI: 10.1083/jcb.200611026
Fibulin-5/DANCE has an elastogenic organizer activity that is abrogated by proteolytic cleavage in vivo
Abstract
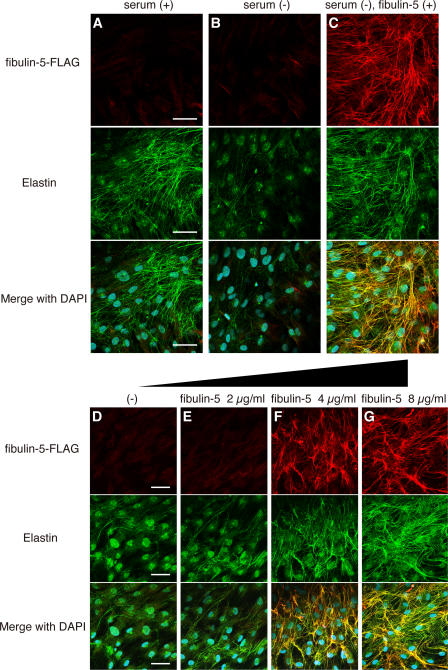
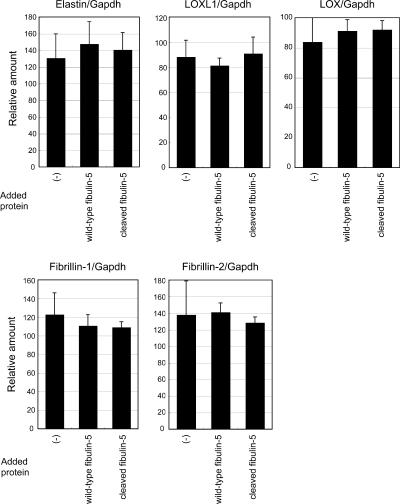
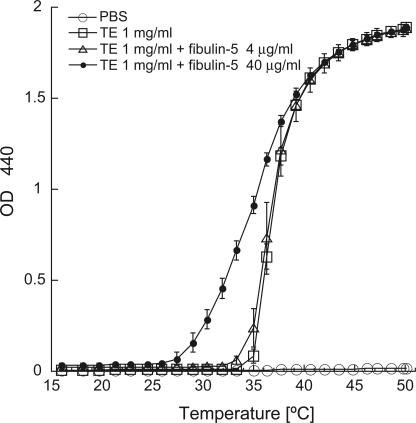
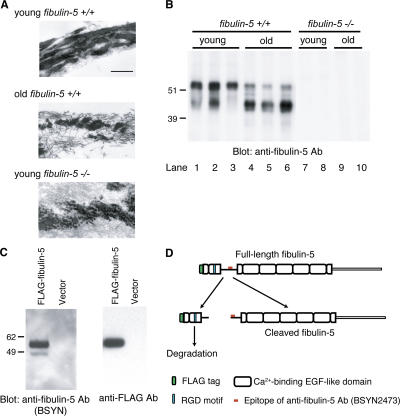
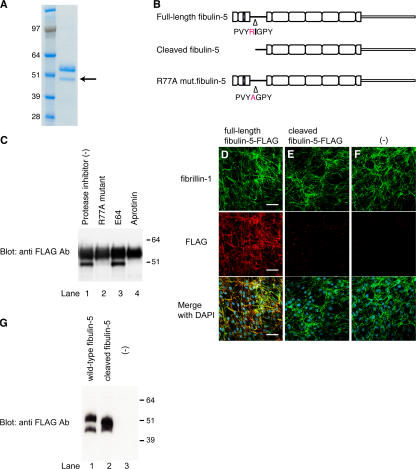
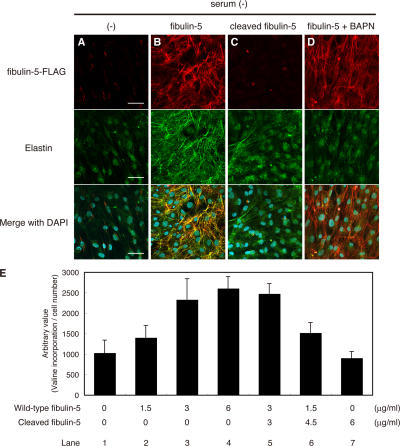
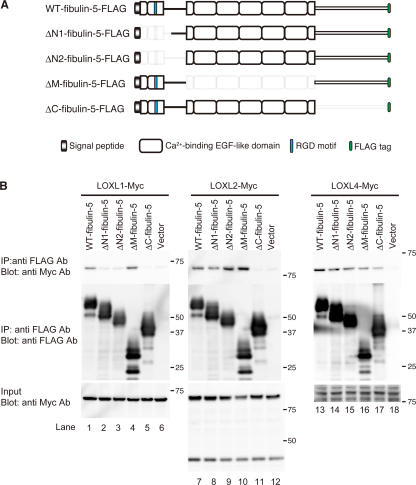
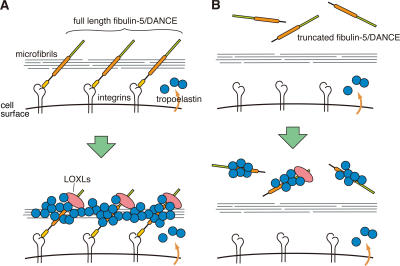
Elastic fibers are required for the elasticity and integrity of various organs. We and others previously showed that fibulin-5 (also called developing arteries and neural crest EGF-like [DANCE] or embryonic vascular EGF-like repeat-containing protein [EVEC]) is indispensable for elastogenesis by studying fibulin-5-deficient mice, which recapitulate human aging phenotypes caused by disorganized elastic fibers (Nakamura, T., P.R. Lozano, Y. Ikeda, Y. Iwanaga, A. Hinek, S. Minamisawa, C.F. Cheng, K. Kobuke, N. Dalton, Y. Takada, et al. 2002. Nature. 415:171-175; Yanagisawa, H., E.C. Davis, B.C. Starcher, T. Ouchi, M. Yanagisawa, J.A. Richardson, and E.N. Olson. 2002. Nature. 415:168-171). However, the molecular mechanism by which fiblin-5 contributes to elastogenesis remains unknown. We report that fibulin-5 protein potently induces elastic fiber assembly and maturation by organizing tropoelastin and cross-linking enzymes onto microfibrils. Deposition of fibulin-5 on microfibrils promotes coacervation and alignment of tropoelastins on microfibrils, and also facilitates cross-linking of tropoelastin by tethering lysyl oxidase-like 1, 2, and 4 enzymes. Notably, recombinant fibulin-5 protein induced elastogenesis even in serum-free conditions, although elastogenesis in cell culture has been believed to be serum-dependent. Moreover, the amount of full-length fibulin-5 diminishes with age, while truncated fibulin-5, which cannot promote elastogenesis, increases. These data suggest that fibulin-5 could be a novel therapeutic target for elastic fiber regeneration.
Figures
References
-
- Bailey, A.J. 2001. Molecular mechanisms of ageing in connective tissues. Mech. Ageing Dev. 122:735–755. - PubMed
-
- Clarke, A.W., S.G. Wise, S.A. Cain, C.M. Kielty, and A.S. Weiss. 2005. Coacervation is promoted by molecular interactions between the PF2 segment of fibrillin-1 and the domain 4 region of tropoelastin. Biochemistry. 44:10271–10281. - PubMed
-
- Clarke, A.W., E.C. Arnspang, S.M. Mithieux, E. Korkmaz, F. Braet, and A.S. Weiss. 2006. Tropoelastin massively associates during coacervation to form quantized protein spheres. Biochemistry. 45:9989–9996. - PubMed
-
- Fisher, G.J., S.C. Datta, H.S. Talwar, Z.Q. Wang, J. Varani, S. Kang, and J.J. Voorhees. 1996. Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature. 379:335–339. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
