

Requirement of the LIM homeodomain transcription factor tailup for normal heart and hematopoietic organ formation in Drosophila melanogaster
- PMID: 17371844
- PMCID: PMC1900034
- DOI: 10.1128/MCB.00093-07
Requirement of the LIM homeodomain transcription factor tailup for normal heart and hematopoietic organ formation in Drosophila melanogaster
Abstract
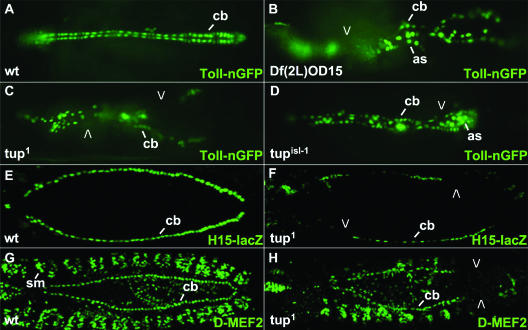
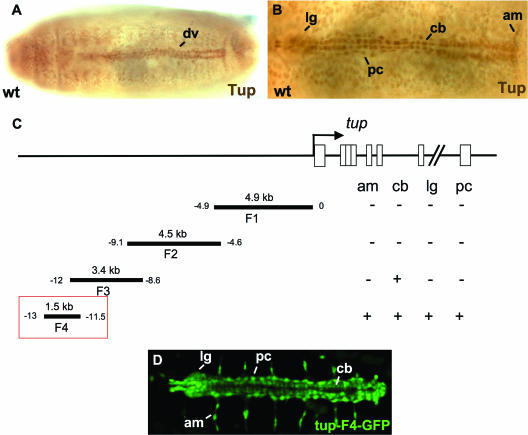
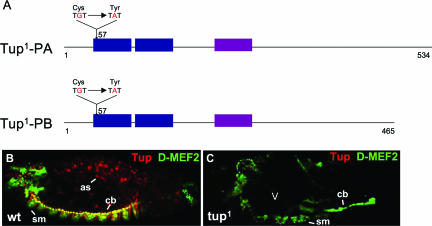
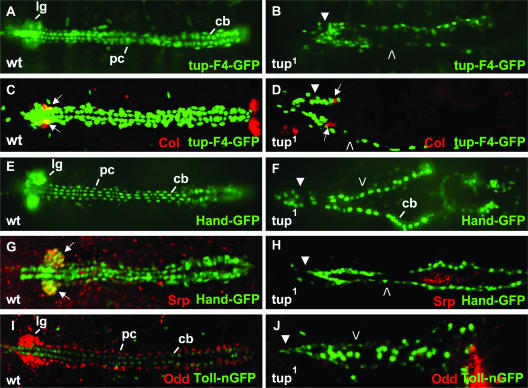
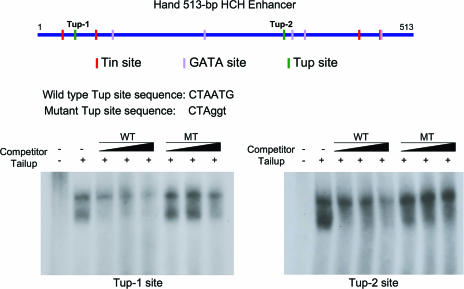
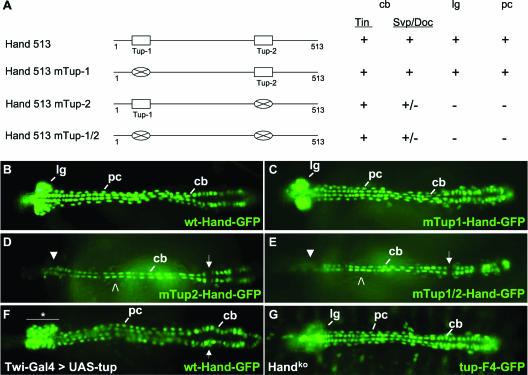
Dorsal vessel morphogenesis in Drosophila melanogaster serves as a superb system with which to study the cellular and genetic bases of heart tube formation. We used a cardioblast-expressed Toll-GFP transgene to screen for additional genes involved in heart development and identified tailup as a locus essential for normal dorsal vessel formation. tailup, related to vertebrate islet1, encodes a LIM homeodomain transcription factor expressed in all cardioblasts and pericardial cells of the heart tube as well as in associated lymph gland hematopoietic organs and alary muscles that attach the dorsal vessel to the epidermis. A transcriptional enhancer regulating expression in these four cell types was identified and used as a tailup-GFP transgene with additional markers to characterize dorsal vessel defects resulting from gene mutations. Two reproducible phenotypes were observed in mutant embryos: hypoplastic heart tubes with misaligned cardioblasts and the absence of most lymph gland and pericardial cells. Conversely, a significant expansion of the lymph glands and abnormal morphology of the heart were observed when tailup was overexpressed in the mesoderm. Tailup was shown to bind to two DNA recognition sequences in the dorsal vessel enhancer of the Hand basic helix-loop-helix transcription factor gene, with one site proven to be essential for the lymph gland, pericardial cell, and Svp/Doc cardioblast expression of Hand. Together, these results establish Tailup as being a critical new transcription factor in dorsal vessel morphogenesis and lymph gland formation and place this regulator directly upstream of Hand in these developmental processes.
Figures

References
-
- Barolo, S., L. A. Carver, and J. W. Posakony. 2000. GFP and β-galactosidase transformation vectors for promoter/enhancer analysis in Drosophila. BioTechniques 29:726-732. - PubMed
-
- Brand, A. H., and N. Perrimon. 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118:401-415. - PubMed
-
- Cripps, R. M., and E. N. Olson. 2002. Control of cardiac development by an evolutionarily conserved transcriptional network. Dev. Biol. 246:14-28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases