

Ubiquitin-independent proteasomal degradation of Fra-1 is antagonized by Erk1/2 pathway-mediated phosphorylation of a unique C-terminal destabilizer
- PMID: 17371847
- PMCID: PMC1900028
- DOI: 10.1128/MCB.01776-06
Ubiquitin-independent proteasomal degradation of Fra-1 is antagonized by Erk1/2 pathway-mediated phosphorylation of a unique C-terminal destabilizer
Abstract
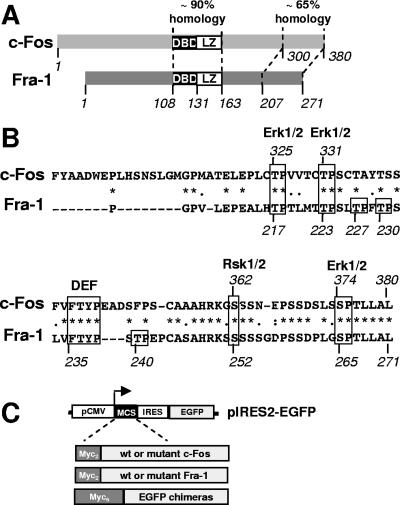
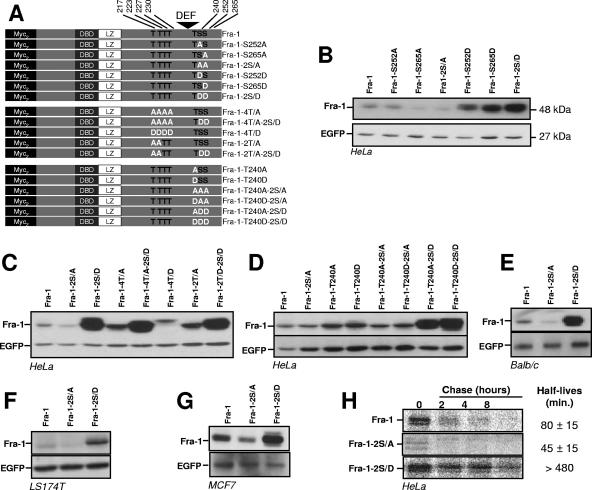
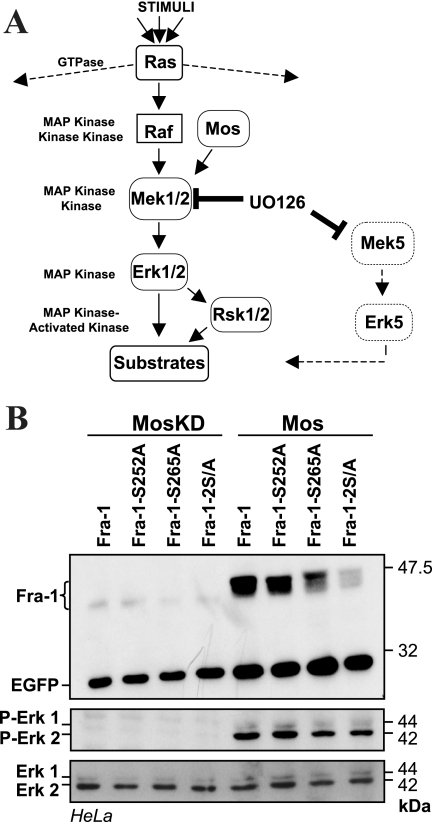
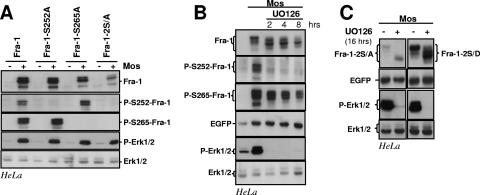
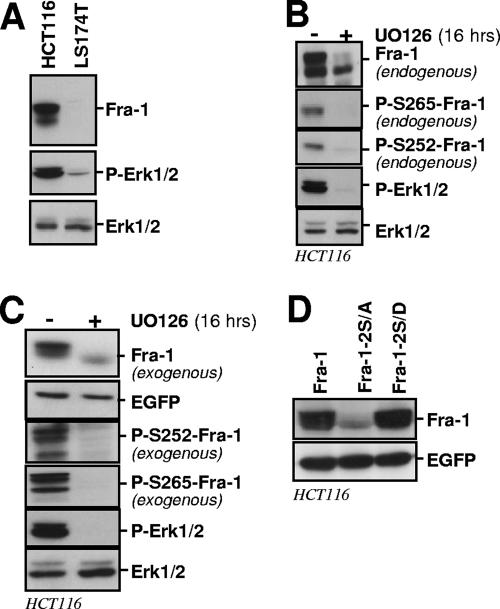
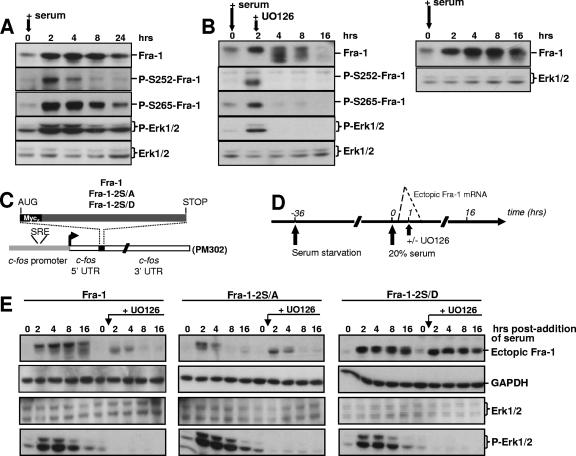
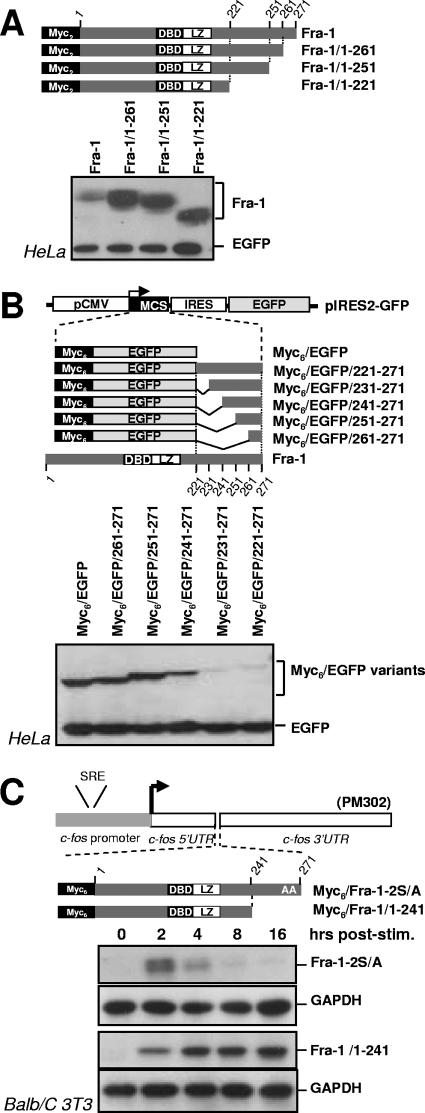
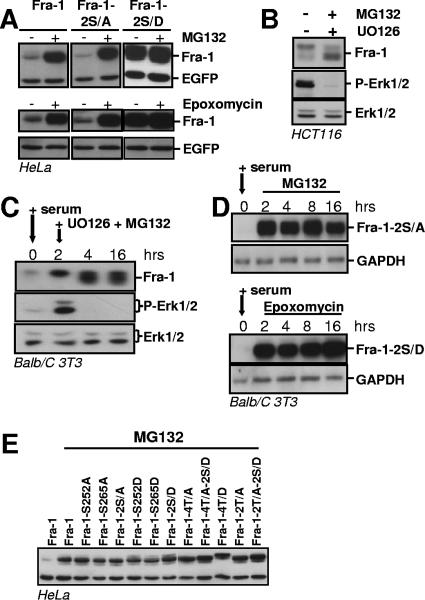
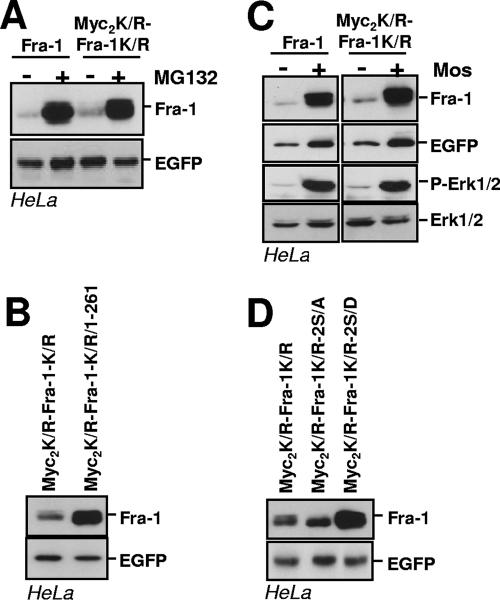
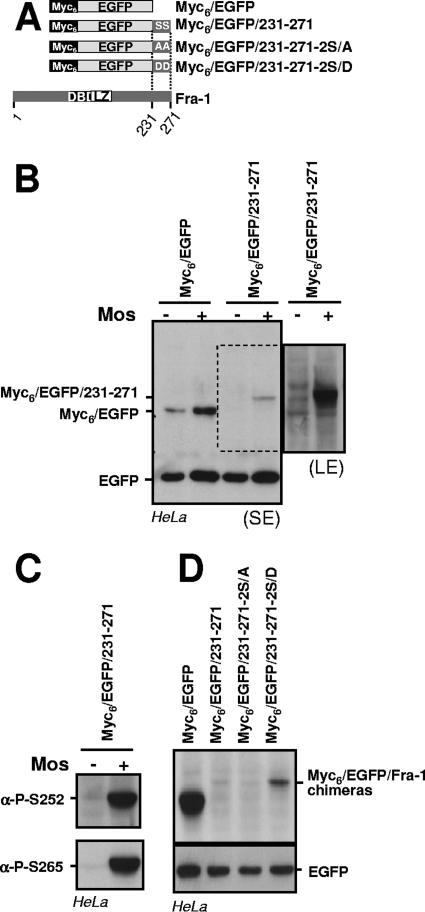
Fra-1, a transcription factor that is phylogenetically and functionally related to the proto-oncoprotein c-Fos, controls many essential cell functions. It is expressed in many cell types, albeit with differing kinetics and abundances. In cells reentering the cell cycle, Fra-1 expression is transiently stimulated albeit later than that of c-Fos and for a longer time. Moreover, Fra-1 overexpression is found in cancer cells displaying high Erk1/2 activity and has been linked to tumorigenesis. One crucial point of regulation of Fra-1 levels is controlled protein degradation, the mechanism of which remains poorly characterized. Here, we have combined genetic, pharmacological, and signaling studies to investigate this process in nontransformed cells and to elucidate how it is altered in cancer cells. We report that the intrinsic instability of Fra-1 depends on a single destabilizer contained within the C-terminal 30 to 40 amino acids. Two serines therein, S252 and S265, are phosphorylated by kinases of the Erk1/2 pathway, which compromises protein destruction upon both normal physiological induction and tumorigenic constitutive activation of this cascade. Our data also indicate that Fra-1, like c-Fos, belongs to a small group of proteins that may, under certain circumstances, undergo ubiquitin-independent degradation by the proteasome. Our work reveals both similitudes and differences between Fra-1 and c-Fos degradation mechanisms. In particular, the presence of a single destabilizer within Fra-1, instead of two that are differentially regulated in c-Fos, explains the much faster turnover of the latter when cells traverse the G(0)/G(1)-to-S-phase transition. Finally, our study offers further insights into the signaling-regulated expression of the other Fos family proteins.
Figures

References
-
- Acquaviva, C., F. Brockly, P. Ferrara, G. Bossis, C. Salvat, I. Jariel-Encontre, and M. Piechaczyk. 2001. Identification of a C-terminal tripeptide motif involved in the control of rapid proteasomal degradation of c-Fos proto-oncoprotein during the G(0)-to-S phase transition. Oncogene 20:7563-7572. - PubMed
-
- Adiseshaiah, P., S. Peddakama, Q. Zhang, D. V. Kalvakolanu, and S. P. Reddy. 2005. Mitogen regulated induction of FRA-1 proto-oncogene is controlled by the transcription factors binding to both serum and TPA response elements. Oncogene 24:4193-4205. - PubMed
-
- Asher, G., Z. Bercovich, P. Tsvetkov, Y. Shaul, and C. Kahana. 2005. 20S proteasomal degradation of ornithine decarboxylase is regulated by NQO1. Mol. Cell 17:645-655. - PubMed
-
- Asher, G., and Y. Shaul. 2005. p53 proteasomal degradation: poly-ubiquitination is not the whole story. Cell Cycle 4:1015-1018. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous