

The role of antibody polyspecificity and lipid reactivity in binding of broadly neutralizing anti-HIV-1 envelope human monoclonal antibodies 2F5 and 4E10 to glycoprotein 41 membrane proximal envelope epitopes
- PMID: 17372000
- PMCID: PMC2262928
- DOI: 10.4049/jimmunol.178.7.4424
The role of antibody polyspecificity and lipid reactivity in binding of broadly neutralizing anti-HIV-1 envelope human monoclonal antibodies 2F5 and 4E10 to glycoprotein 41 membrane proximal envelope epitopes
Abstract
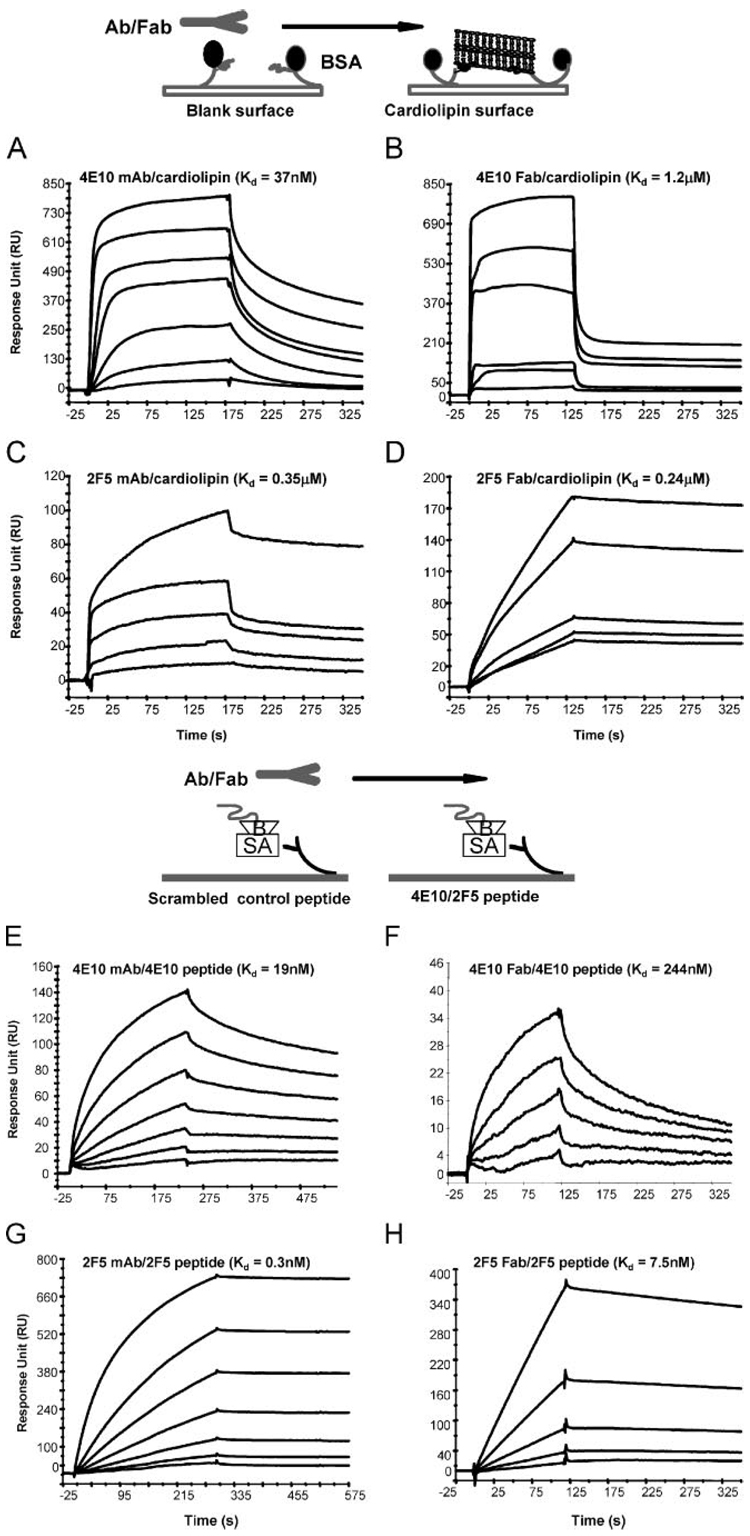
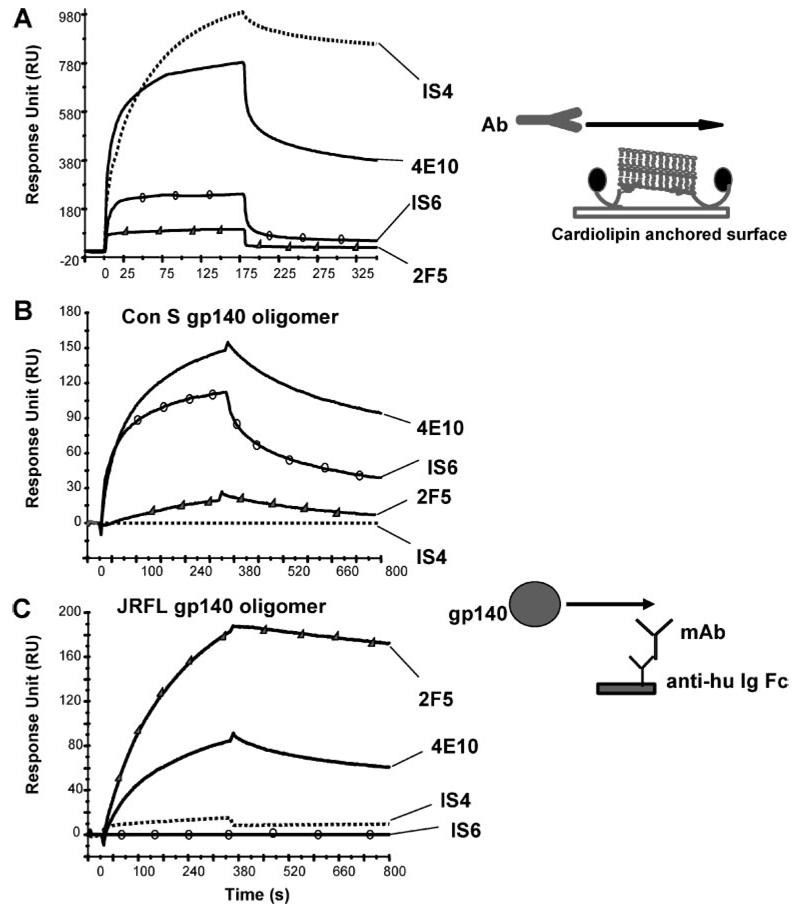
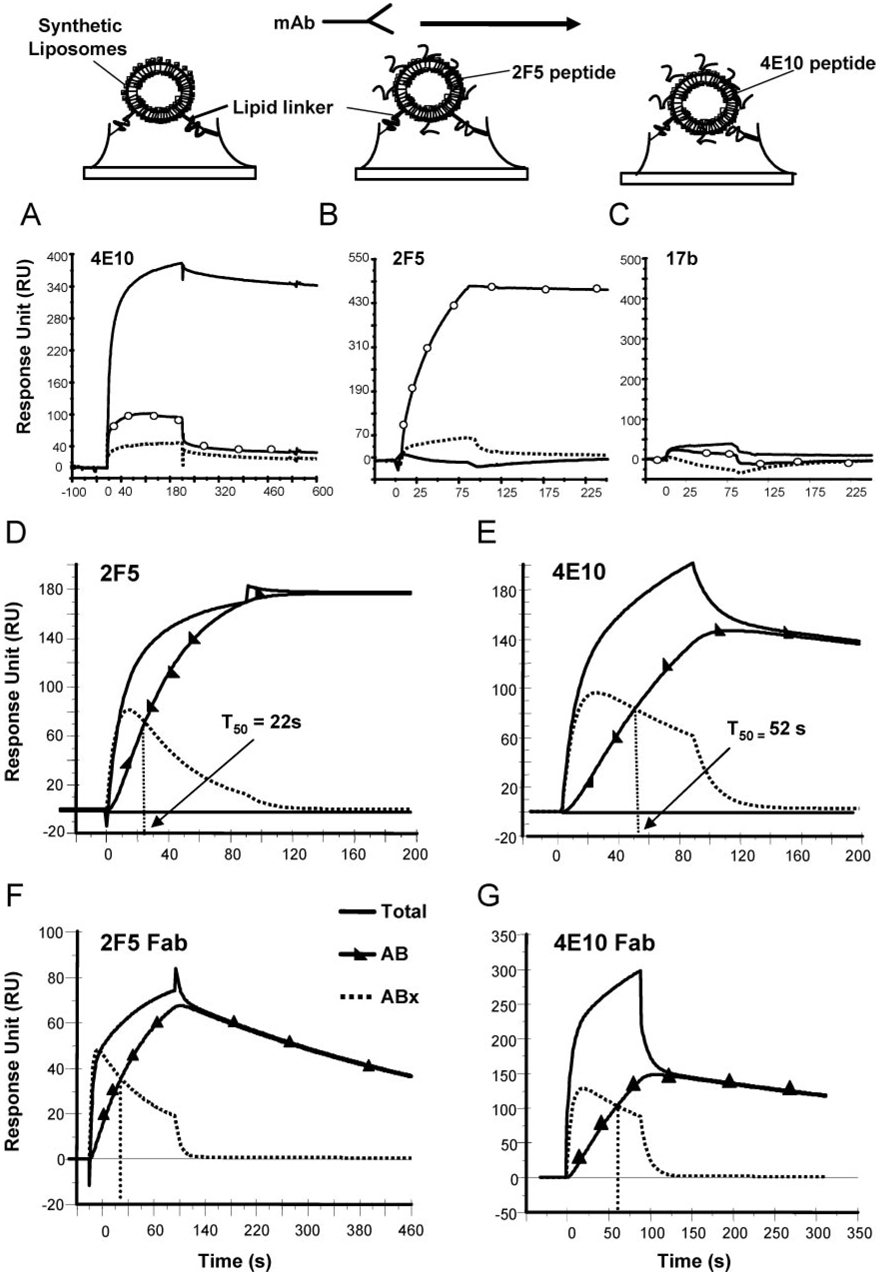
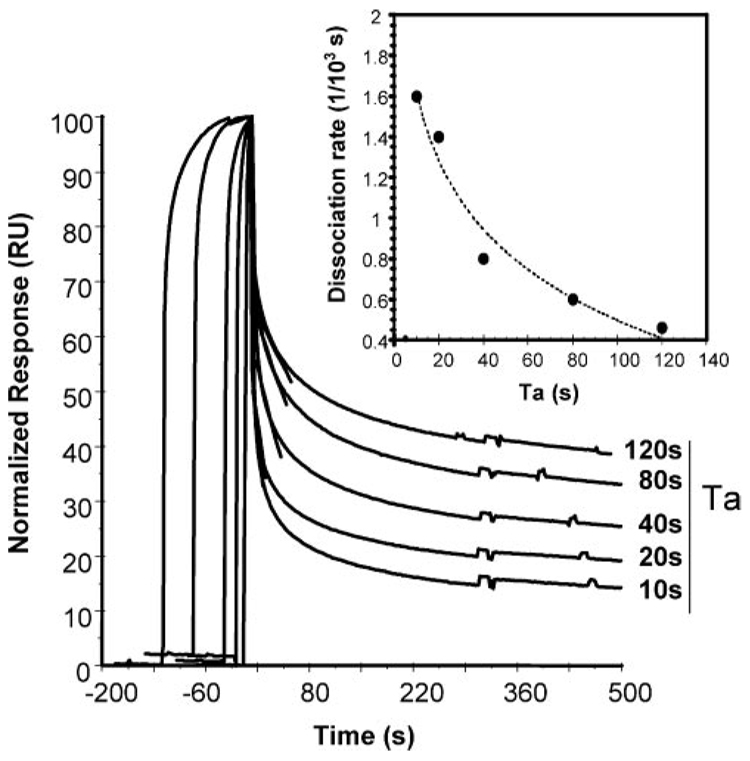
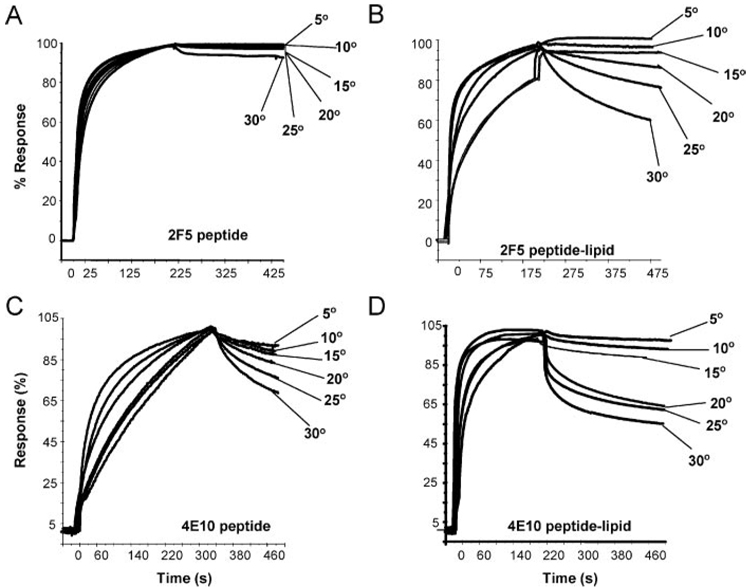
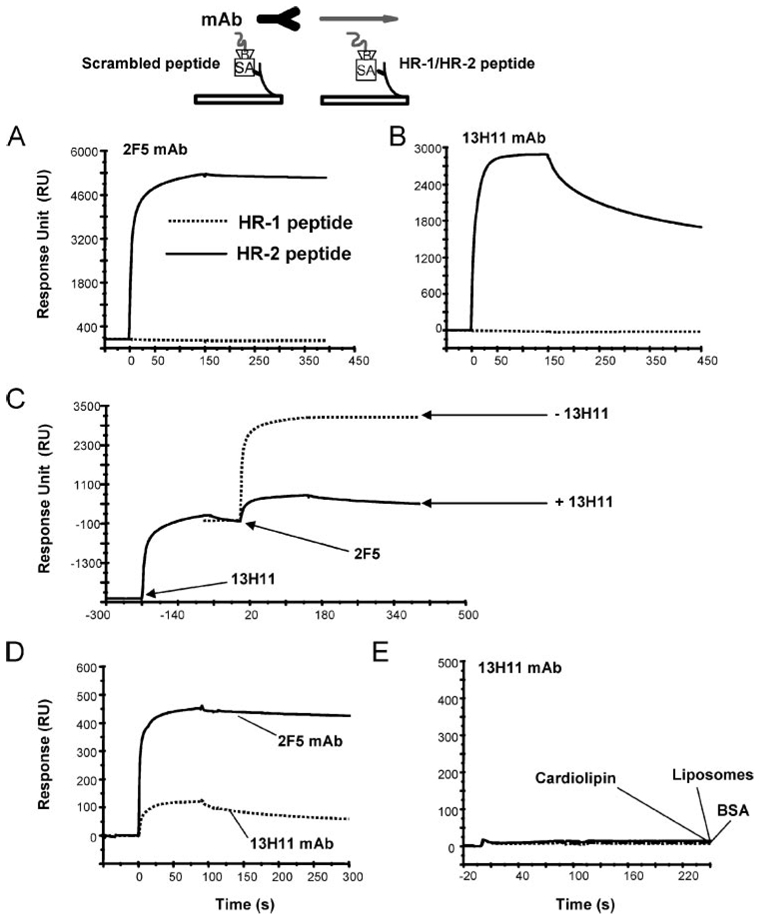
Two neutralizing human mAbs, 2F5 and 4E10, that react with the HIV-1 envelope gp41 membrane proximal region are also polyspecific autoantibodies that bind to anionic phospholipids. To determine the autoantibody nature of these Abs, we have compared their reactivities with human anti-cardiolipin mAbs derived from a primary antiphospholipid syndrome patient. To define the role of lipid polyreactivity in binding of 2F5 and 4E10 mAbs to HIV-1 envelope membrane proximal epitopes, we determined the kinetics of binding of mAbs 2F5 and 4E10 to their nominal gp41 epitopes vs liposome-gp41 peptide conjugates. Both anti-HIV-1 mAbs 2F5 and 4E10 bound to cardiolipin with K(d) values similar to those of autoimmune anti-cardiolipin Abs, IS4 and IS6. Binding kinetics studies revealed that mAb 2F5 and 4E10 binding to their respective gp41 peptide-lipid conjugates could best be defined by a two-step (encounter-docking) conformational change model. In contrast, binding of 2F5 and 4E10 mAbs to linear peptide epitopes followed a simple Langmuir model. A mouse mAb, 13H11, that cross-blocks mAb 2F5 binding to the gp41 epitope did not cross-react with lipids nor did it neutralize HIV-1 viruses. Taken together, these data demonstrate the similarity of 2F5 and 4E10 mAbs to known anti-cardiolipin Abs and support the model that mAb 2F5 and 4E10 binding to HIV-1 involves both viral lipid membrane and gp41 membrane proximal epitopes.
Conflict of interest statement
Figures
References
-
- Cardoso RM, Zwick MB, Stanfield RL, Kunert R, Binley JM, Katinger H, Burton DR, Wilson IA. Broadly neutralizing anti-HIV antibody 4E10 recognizes a helical conformation of a highly conserved fusion-associated motif in gp41. Immunity. 2005;22:163–173. - PubMed
-
- Stiegler G, Kunert R, Purtscher M, Wolbank S, Voglauer R, Steindl F, Katinger H. A potent cross-clade neutralizing human monoclonal antibody against a novel epitope on gp41 of human immunodeficiency virus type 1. AIDS Res. Hum. Retroviruses. 2001;17:1757–1765. - PubMed
-
- D’Souza MP, Livnat D, Bradac JA, Bridges SH. Evaluation of monoclonal antibodies to human immunodeficiency virus type 1 primary isolates by neutralization assays: performance criteria for selecting candidate antibodies for clinical trials. AIDS Clinical Trials Group Antibody Selection Working Group. J. Infect. Dis. 1997;175:1056–1062. - PubMed
-
- Barbato GE, Baianchi P, Inagallinella P, Hurni WH, Miller MD, Ciliberto R, Cortrese R, Bazzo R, Shiver JW, Pessi A. Structural analysis of the epitope of the anti-HIV antibody 2F5 sheds light into its mechanism of neutralization and HIV fusion. J. Mol. Biol. 2003;330:1101–1115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
