

Normal development and function of invariant natural killer T cells in mice with isoglobotrihexosylceramide (iGb3) deficiency
- PMID: 17372206
- PMCID: PMC1851602
- DOI: 10.1073/pnas.0611139104
Normal development and function of invariant natural killer T cells in mice with isoglobotrihexosylceramide (iGb3) deficiency
Abstract
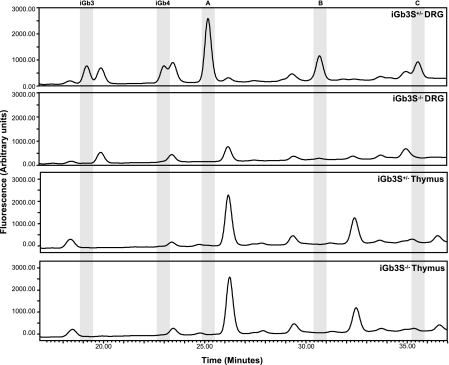
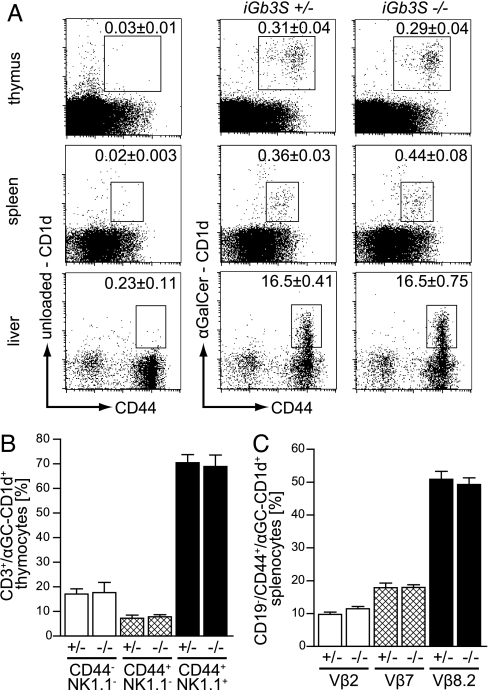
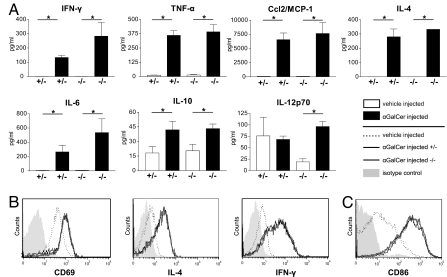
CD1d-restricted natural killer T (NKT) cells, expressing the invariant T cell antigen receptor (TCR) chain encoded by Valpha14-Jalpha18 gene segments in mice and Valpha24-Jalpha18 in humans [invariant NKT (iNKT) cells], contribute to immunoregulatory processes, such as tolerance, host defense, and tumor surveillance. iNKT cells are positively selected in the thymus by CD1d molecules expressed by CD4(+)/CD8(+) cortical thymocytes. However, the identity of the endogenous lipid(s) responsible for positive selection of iNKT cells remains unclear. One candidate lipid proposed to play a role in positive selection is isoglobotrihexosylceramide (iGb3). However, no direct evidence for its physiological role has been provided. Therefore, to directly investigate the role of iGb3 in iNKT cell selection, we have generated mice deficient in iGb3 synthase [iGb3S, also known as alpha1-3galactosyltransferase 2 (A3galt2)]. These mice developed, grew, and reproduced normally and exhibited no overt behavioral abnormalities. Consistent with the notion that iGb3 is synthesized only by iGb3S, lack of iGb3 in the dorsal root ganglia of iGb3S-deficient mice (iGb3S(-/-)), as compared with iGb3S(+/-) mice, was confirmed. iGb3S(-/-) mice showed normal numbers of iNKT cells in the thymus, spleen, and liver with selected TCR Vbeta chains identical to controls. Upon administration of alpha-galactosylceramide, activation of iNKT and dendritic cells was similar in iGb3S(-/-) and iGb3S(+/-) mice, as measured by up-regulation of CD69 as well as intracellular IL-4 and IFN-gamma in iNKT cells, up-regulation of CD86 on dendritic cells, and rise in serum concentrations of IL-4, IL-6, IL-10, IL-12p70, IFN-gamma, TNF-alpha, and Ccl2/MCP-1. Our results strongly suggest that iGb3 is unlikely to be an endogenous CD1d lipid ligand determining thymic iNKT selection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Natural killer T cells: know thyself.Proc Natl Acad Sci U S A. 2007 Apr 3;104(14):5713-4. doi: 10.1073/pnas.0701493104. Epub 2007 Mar 27. Proc Natl Acad Sci U S A. 2007. PMID: 17389403 Free PMC article. No abstract available.
Similar articles
-
Humans lack iGb3 due to the absence of functional iGb3-synthase: implications for NKT cell development and transplantation.PLoS Biol. 2008 Jul 15;6(7):e172. doi: 10.1371/journal.pbio.0060172. PLoS Biol. 2008. PMID: 18630988 Free PMC article.
-
Globosides but not isoglobosides can impact the development of invariant NKT cells and their interaction with dendritic cells.J Immunol. 2012 Sep 15;189(6):3007-17. doi: 10.4049/jimmunol.1201483. Epub 2012 Aug 8. J Immunol. 2012. PMID: 22875802 Free PMC article.
-
Implications for invariant natural killer T cell ligands due to the restricted presence of isoglobotrihexosylceramide in mammals.Proc Natl Acad Sci U S A. 2007 Apr 3;104(14):5971-6. doi: 10.1073/pnas.0607285104. Epub 2007 Mar 19. Proc Natl Acad Sci U S A. 2007. PMID: 17372214 Free PMC article.
-
[iNKT cells, a friend or a foe for autoimmune disease and allergy?].Nihon Rinsho Meneki Gakkai Kaishi. 2006 Feb;29(1):27-36. doi: 10.2177/jsci.29.27. Nihon Rinsho Meneki Gakkai Kaishi. 2006. PMID: 16505600 Review. Japanese.
-
Functions of CD1d-Restricted Invariant Natural Killer T Cells in Antimicrobial Immunity and Potential Applications for Infection Control.Front Immunol. 2018 Jun 6;9:1266. doi: 10.3389/fimmu.2018.01266. eCollection 2018. Front Immunol. 2018. PMID: 29928278 Free PMC article. Review.
Cited by
-
Invariant natural killer T cells in adipose tissue: novel regulators of immune-mediated metabolic disease.Cell Mol Life Sci. 2013 Dec;70(24):4711-27. doi: 10.1007/s00018-013-1414-1. Epub 2013 Jul 9. Cell Mol Life Sci. 2013. PMID: 23835837 Free PMC article. Review.
-
Thymic development of unconventional T cells: how NKT cells, MAIT cells and γδ T cells emerge.Nat Rev Immunol. 2020 Dec;20(12):756-770. doi: 10.1038/s41577-020-0345-y. Epub 2020 Jun 24. Nat Rev Immunol. 2020. PMID: 32581346 Review.
-
The ins and outs of type I iNKT cell development.Mol Immunol. 2019 Jan;105:116-130. doi: 10.1016/j.molimm.2018.09.023. Epub 2018 Nov 28. Mol Immunol. 2019. PMID: 30502719 Free PMC article. Review.
-
The effect of intracellular trafficking of CD1d on the formation of TCR repertoire of NKT cells.BMB Rep. 2014 May;47(5):241-8. doi: 10.5483/bmbrep.2014.47.5.077. BMB Rep. 2014. PMID: 24755556 Free PMC article. Review.
-
Antigen specificity of invariant natural killer T-cells.Biomed J. 2015 Dec;38(6):470-83. doi: 10.1016/j.bj.2016.01.003. Epub 2016 Mar 10. Biomed J. 2015. PMID: 27013447 Free PMC article. Review.
References
-
- Kasahara M, Nakaya J, Satta Y, Takahata N. Trends Genet. 1997;13:90–92. - PubMed
-
- Brigl M, Brenner MB. Annu Rev Immunol. 2004;22:817–890. - PubMed
-
- Porcelli SA, Modlin RL. Annu Rev Immunol. 1999;17:297–329. - PubMed
-
- Kronenberg M, Gapin L. Nat Rev Immunol. 2002;2:557–568. - PubMed
-
- Makino Y, Kanno R, Ito T, Higashino K, Taniguchi M. Int Immunol. 1995;7:1157–1161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
