

Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex
- PMID: 17372627
- PMCID: PMC1810431
- DOI: 10.1371/journal.pone.0000288
Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex
Abstract
Invertebrates have contributed greatly to our understanding of associative learning because they allow learning protocols to be combined with experimental access to the nervous system. The honeybee Apis mellifera constitutes a standard model for the study of appetitive learning and memory since it was shown, almost a century ago, that bees learn to associate different sensory cues with a reward of sugar solution. However, up to now, no study has explored aversive learning in bees in such a way that simultaneous access to its neural bases is granted. Using odorants paired with electric shocks, we conditioned the sting extension reflex, which is exhibited by harnessed bees when subjected to a noxious stimulation. We show that this response can be conditioned so that bees learn to extend their sting in response to the odorant previously punished. Bees also learn to extend the proboscis to one odorant paired with sugar solution and the sting to a different odorant paired with electric shock, thus showing that they can master both appetitive and aversive associations simultaneously. Responding to the appropriate odorant with the appropriate response is possible because two different biogenic amines, octopamine and dopamine subserve appetitive and aversive reinforcement, respectively. While octopamine has been previously shown to substitute for appetitive reinforcement, we demonstrate that blocking of dopaminergic, but not octopaminergic, receptors suppresses aversive learning. Therefore, aversive learning in honeybees can now be accessed both at the behavioral and neural levels, thus opening new research avenues for understanding basic mechanisms of learning and memory.
Conflict of interest statement
Figures

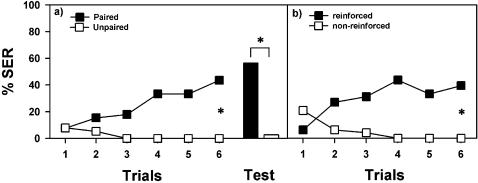





References
-
- Menzel R. Memory dynamics in the honeybee. J Comp Physiol A. 1999;185:323–340.
-
- Menzel R. Searching for the memory trace in a mini-brain, the honeybee. Learn Mem. 2001;8:53–62. - PubMed
-
- Menzel R, Giurfa M. Cognitive architecture of a mini-brain: the honeybee. Trends Cogn Sci. 2001;5:62–71. - PubMed
-
- Giurfa M. Cognitive neuroethology: dissecting non-elemental learning in a honeybee brain. Curr Opin Neurobiol. 2003;13:R726–R735. - PubMed
-
- Frisch K von. The Dance Language and Orientation of Bees. Cambridge: Harvard University Press; 1967. p. 566 p.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
