

The making of a social insect: developmental architectures of social design
- PMID: 17373656
- PMCID: PMC2398704
- DOI: 10.1002/bies.20549
The making of a social insect: developmental architectures of social design
Abstract
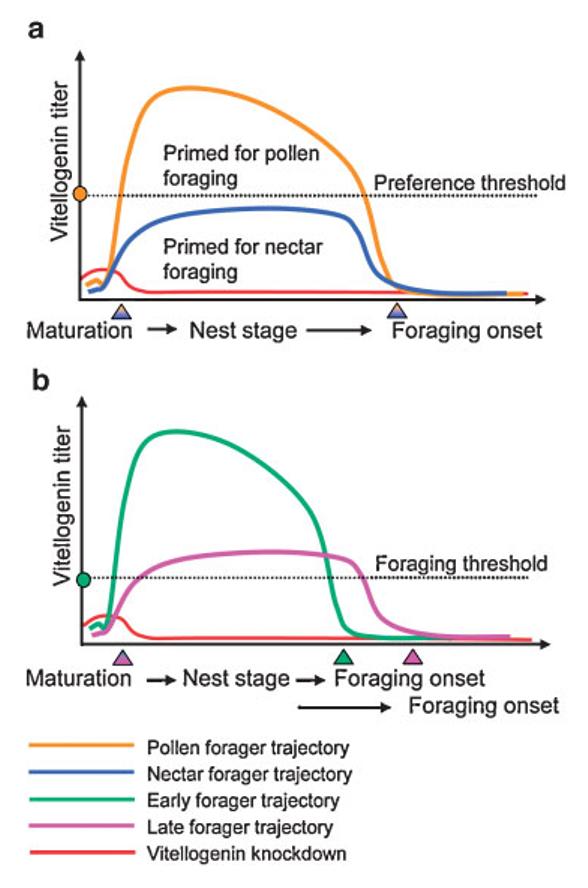
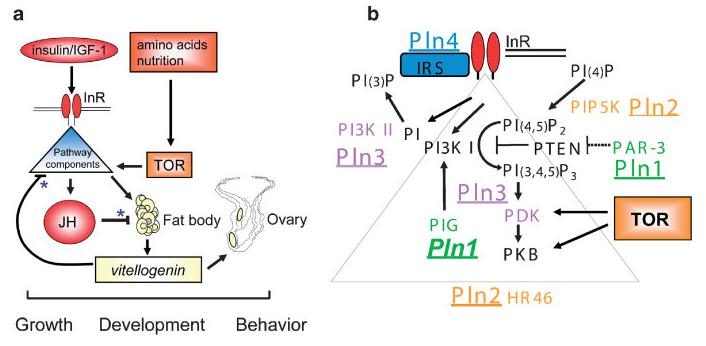
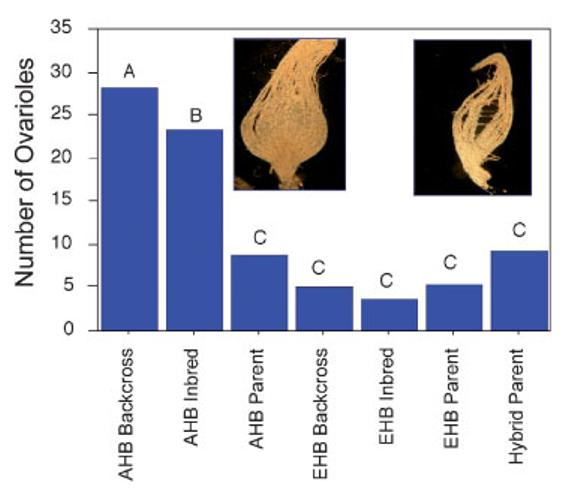
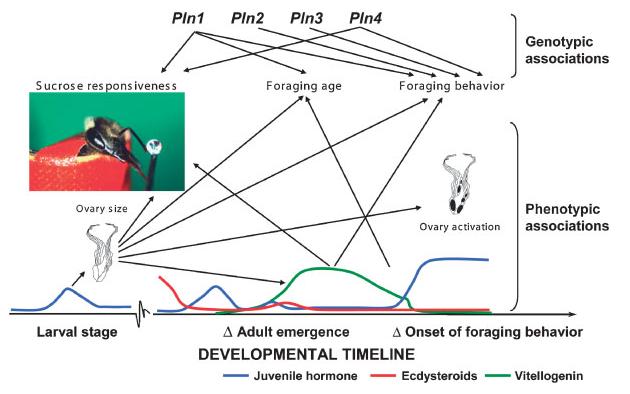
We marvel at the social complexity of insects, marked by anatomically and behaviorally distinguishable castes, division of labor and specialization--but how do such systems evolve? Insect societies are composed of individuals, each undergoing its own developmental process and each containing its own genetic information and experiencing its own developmental and experiential environment. Yet societies appear to function as if the colonies themselves are individuals with novel "social genes" and novel social developmental processes. We propose an alternative hypothesis. The origins of complex social behavior, from which insect societies emerge, are derived from ancestral developmental programs. These programs originated in ancient solitary insects and required little evolutionary remodeling. We present evidence from behavioral assays, selective breeding, genetic mapping, functional genomics and endocrinology, and comparative anatomy and physiology. These insights explain how complex social behavior can evolve from heterochronic changes in reproductive signaling systems that govern ubiquitous and ancient relationships between behavior and ovarian development.
(c) 2007 Wiley Periodicals, Inc.
Figures

References
-
- Winston ML. The Biology of the Honey Bee. Harvard University Press; Cambridge MA: 1987.
-
- Snodgrass RE. Anatomy of the Honey Bee. Comstock Publishing Associates; New York: 1956.
-
- Evans JD, Wheeler DE. Gene expression and the evolution of insect polyphenisms. Bioessays. 2001;23:62–68. - PubMed
-
- Bishop GH. Fertilization in the honeybee. J Exp Zool. 1920;31:225–286.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
