

H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state
- PMID: 17373856
- PMCID: PMC1828143
- DOI: 10.1371/journal.pbio.0050081
H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state
Abstract
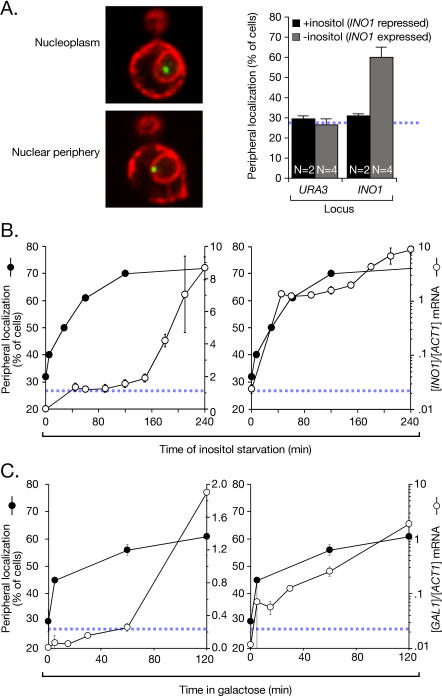
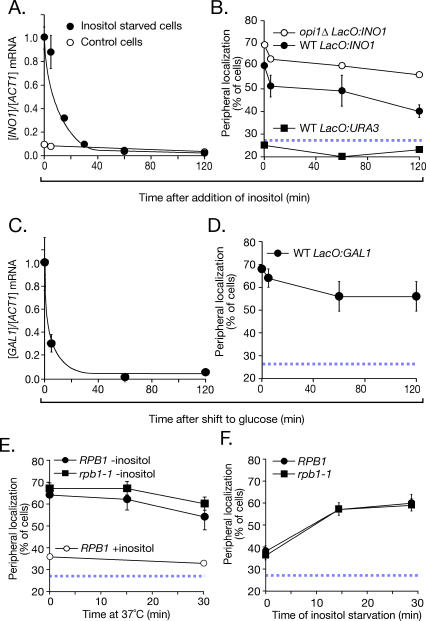
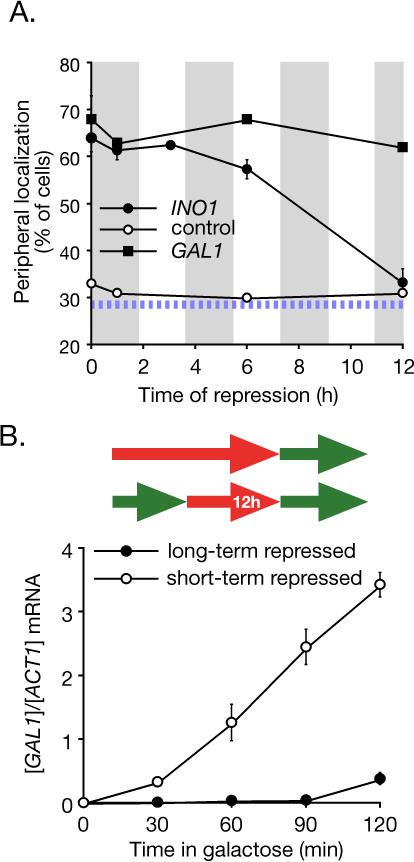
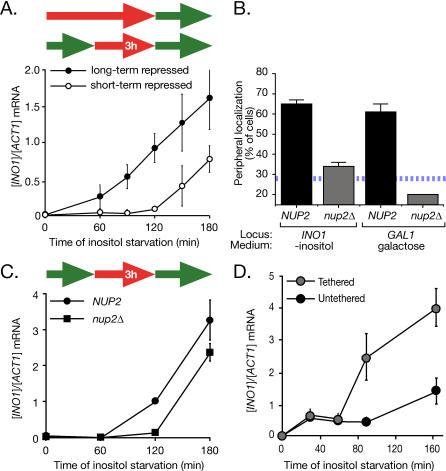
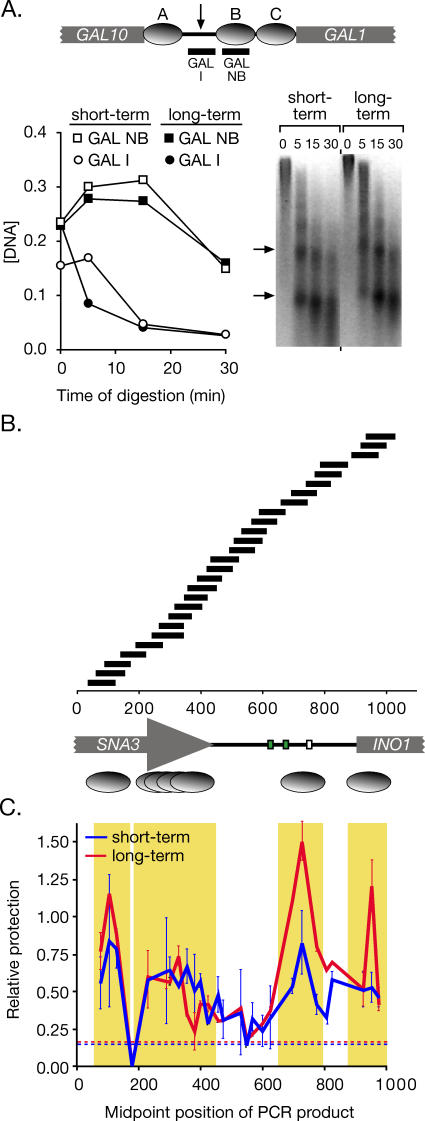
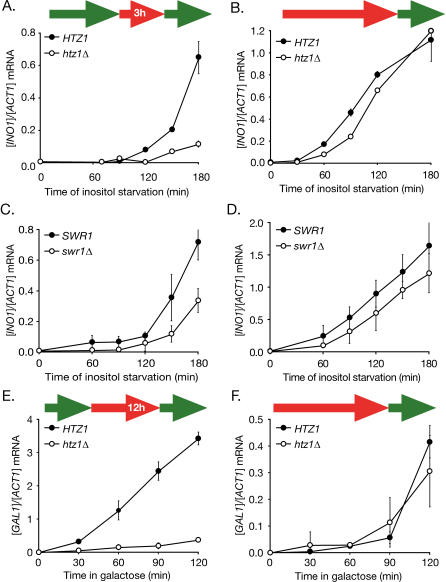
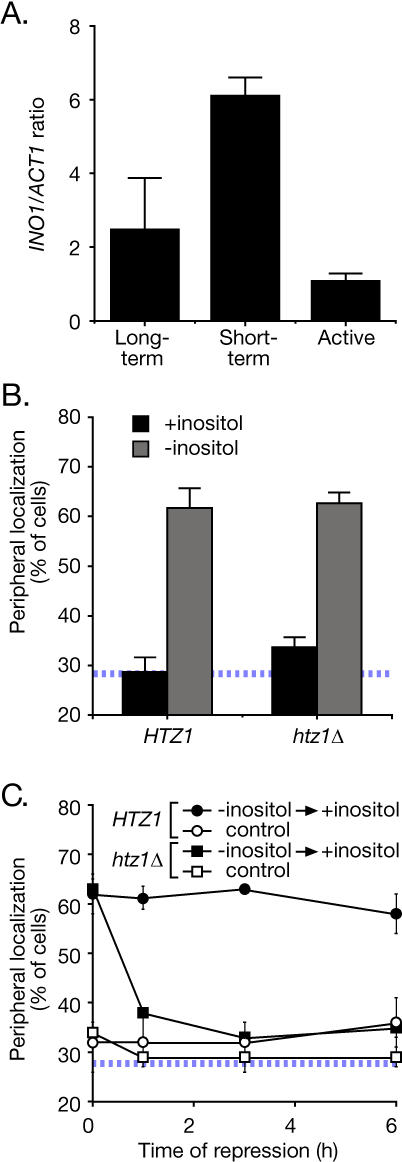
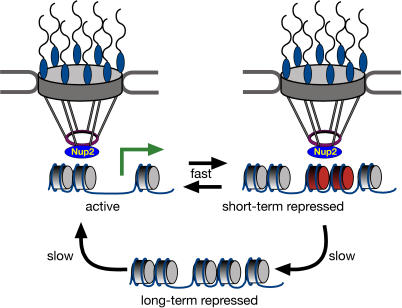
Many genes are recruited to the nuclear periphery upon transcriptional activation. The mechanism and functional significance of this recruitment is unclear. We find that recruitment of the yeast INO1 and GAL1 genes to the nuclear periphery is rapid and independent of transcription. Surprisingly, these genes remain at the periphery for generations after they are repressed. Localization at the nuclear periphery serves as a form of memory of recent transcriptional activation, promoting reactivation. Previously expressed GAL1 at the nuclear periphery is activated much more rapidly than long-term repressed GAL1 in the nucleoplasm, even after six generations of repression. Localization of INO1 at the nuclear periphery is necessary and sufficient to promote more rapid activation. This form of transcriptional memory is chromatin based; the histone variant H2A.Z is incorporated into nucleosomes within the recently repressed INO1 promoter and is specifically required for rapid reactivation of both INO1 and GAL1. Furthermore, H2A.Z is required to retain INO1 at the nuclear periphery after repression. Therefore, H2A.Z-mediated localization of recently repressed genes at the nuclear periphery represents an epigenetic state that confers memory of transcriptional activation and promotes reactivation.
Conflict of interest statement
Figures
References
-
- Cockell M, Gasser SM. Nuclear compartments and gene regulation. Curr Opin Genet Dev. 1999;9:199–205. - PubMed
-
- Isogai Y, Tjian R. Targeting genes and transcription factors to segregated nuclear compartments. Curr Opin Cell Biol. 2003;15:296–303. - PubMed
-
- Brown KE, Guest SS, Smale ST, Hahm K, Merkenschlager M, et al. Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell. 1997;91:845–854. - PubMed
-
- Csink AK, Henikoff S. Genetic modification of heterochromatic association and nuclear organization in Drosophila . Nature. 1996;381:529–531. - PubMed
-
- Dernburg AF, Broman KW, Fung JC, Marshall WF, Philips J, et al. Perturbation of nuclear architecture by long-distance chromosome interactions. Cell. 1996;85:745–759. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
