

Protein oxidation implicated as the primary determinant of bacterial radioresistance
- PMID: 17373858
- PMCID: PMC1828145
- DOI: 10.1371/journal.pbio.0050092
Protein oxidation implicated as the primary determinant of bacterial radioresistance
Abstract
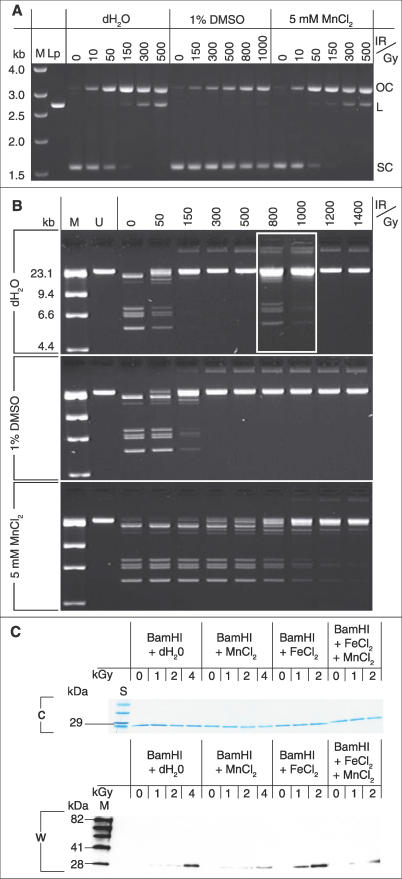
In the hierarchy of cellular targets damaged by ionizing radiation (IR), classical models of radiation toxicity place DNA at the top. Yet, many prokaryotes are killed by doses of IR that cause little DNA damage. Here we have probed the nature of Mn-facilitated IR resistance in Deinococcus radiodurans, which together with other extremely IR-resistant bacteria have high intracellular Mn/Fe concentration ratios compared to IR-sensitive bacteria. For in vitro and in vivo irradiation, we demonstrate a mechanistic link between Mn(II) ions and protection of proteins from oxidative modifications that introduce carbonyl groups. Conditions that inhibited Mn accumulation or Mn redox cycling rendered D. radiodurans radiation sensitive and highly susceptible to protein oxidation. X-ray fluorescence microprobe analysis showed that Mn is globally distributed in D. radiodurans, but Fe is sequestered in a region between dividing cells. For a group of phylogenetically diverse IR-resistant and IR-sensitive wild-type bacteria, our findings support the idea that the degree of resistance is determined by the level of oxidative protein damage caused during irradiation. We present the case that protein, rather than DNA, is the principal target of the biological action of IR in sensitive bacteria, and extreme resistance in Mn-accumulating bacteria is based on protein protection.
Conflict of interest statement
Figures





Comment in
-
Paradox resolved? The strange case of the radiation-resistant bacteria.PLoS Biol. 2007 Apr;5(4):e108. doi: 10.1371/journal.pbio.0050108. Epub 2007 Mar 20. PLoS Biol. 2007. PMID: 20076666 Free PMC article. No abstract available.
References
-
- Daly MJ, Gaidamakova EK, Matrosova VY, Vasilenko A, Zhai M, et al. Accumulation of Mn(II) in Deinococcus radiodurans facilitates gamma-radiation resistance. Science. 2004;306:1025–1028. - PubMed
-
- Gerard E, Jolivet E, Prieur D, Forterre P. DNA protection mechanisms are not involved in the radioresistance of the hyperthermophilic archaea Pyrococcus abyssi and P. furiosus . Mol Genet Genomics. 2001;266:72–78. - PubMed
-
- Cox MM, Battista JR. Deinococcus radiodurans—The consummate survivor. Nat Rev Microbiol. 2005;3:882–892. - PubMed
-
- Ghosal D, Omelchenko MV, Gaidamakova EK, Matrosova VY, Vasilenko A, et al. How radiation kills cells: Survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. FEMS Microbiol Rev. 2005;29:361–375. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
