

Crystal structure of the carbapenemase OXA-24 reveals insights into the mechanism of carbapenem hydrolysis
- PMID: 17374723
- PMCID: PMC1838445
- DOI: 10.1073/pnas.0607557104
Crystal structure of the carbapenemase OXA-24 reveals insights into the mechanism of carbapenem hydrolysis
Abstract
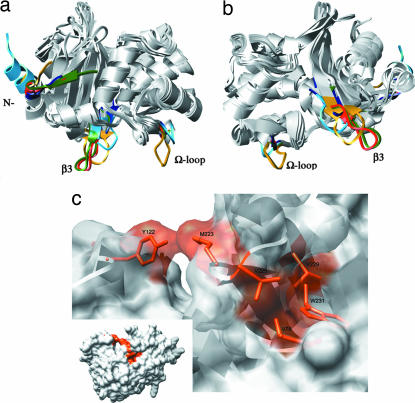
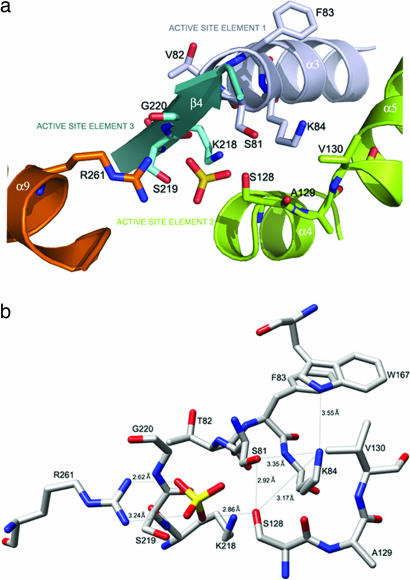
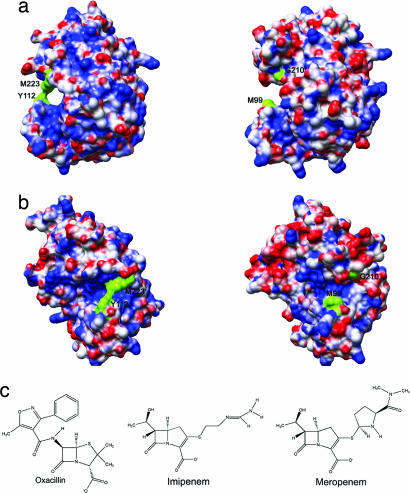
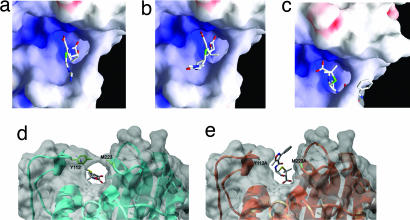
Combating bacterial resistance to beta-lactams, the most widely used antibiotics, is an emergent and clinically important challenge. OXA-24 is a class D beta-lactamase isolated from a multiresistant epidemic clinical strain of Acinetobacter baumannii. We have investigated how OXA-24 specifically hydrolyzes the last resort carbapenem antibiotic, and we have determined the crystal structure of OXA-24 at a resolution of 2.5 A. The structure shows that the carbapenem's substrate specificity is determined by a hydrophobic barrier that is established through the specific arrangement of the Tyr-112 and Met-223 side chains, which define a tunnel-like entrance to the active site. The importance of these residues was further confirmed by mutagenesis studies. Biochemical and microbiological analyses of specific point mutants selected on the basis of structural criteria significantly reduced the catalytic efficiency (k(cat)/K(m)) against carbapenems, whereas the specificity for oxacillin was noticeably increased. This is the previously unrecognized crystal structure that has been obtained for a class D carbapenemase enzyme. Accordingly, this information may help to improve the development of effective new drugs to combat beta-lactam resistance. More specifically, it may help to overcome carbapenem resistance in A. baumannii, probably one of the most worrying infectious threats in hospitals worldwide.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Crystal structure of the OXA-48 beta-lactamase reveals mechanistic diversity among class D carbapenemases.Chem Biol. 2009 May 29;16(5):540-7. doi: 10.1016/j.chembiol.2009.04.010. Chem Biol. 2009. PMID: 19477418
-
Structure of ADC-68, a novel carbapenem-hydrolyzing class C extended-spectrum β-lactamase isolated from Acinetobacter baumannii.Acta Crystallogr D Biol Crystallogr. 2014 Nov;70(Pt 11):2924-36. doi: 10.1107/S1399004714019543. Epub 2014 Oct 23. Acta Crystallogr D Biol Crystallogr. 2014. PMID: 25372683
-
Conformational stability of OXA-51 β-lactamase explains its role in carbapenem resistance of Acinetobacter baumannii.J Biomol Struct Dyn. 2014;32(9):1406-20. doi: 10.1080/07391102.2013.819789. Epub 2013 Jul 24. J Biomol Struct Dyn. 2014. PMID: 23879430
-
Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology.Clin Microbiol Infect. 2006 Sep;12(9):826-36. doi: 10.1111/j.1469-0691.2006.01456.x. Clin Microbiol Infect. 2006. PMID: 16882287 Review.
-
Decoding the Structural Basis For Carbapenem Hydrolysis By Class A β-lactamases: Fishing For A Pharmacophore.Curr Drug Targets. 2016;17(9):983-1005. doi: 10.2174/1389450116666151001104448. Curr Drug Targets. 2016. PMID: 26424401 Review.
Cited by
-
Structural basis of activity against aztreonam and extended spectrum cephalosporins for two carbapenem-hydrolyzing class D β-lactamases from Acinetobacter baumannii.Biochemistry. 2015 Mar 17;54(10):1976-87. doi: 10.1021/bi501547k. Epub 2015 Mar 2. Biochemistry. 2015. PMID: 25710192 Free PMC article.
-
Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace.Biomolecules. 2020 May 6;10(5):720. doi: 10.3390/biom10050720. Biomolecules. 2020. PMID: 32384624 Free PMC article. Review.
-
Class D β-lactamases: are they all carbapenemases?Antimicrob Agents Chemother. 2014;58(4):2119-25. doi: 10.1128/AAC.02522-13. Epub 2014 Jan 27. Antimicrob Agents Chemother. 2014. PMID: 24468778 Free PMC article.
-
Identifying Oxacillinase-48 Carbapenemase Inhibitors Using DNA-Encoded Chemical Libraries.ACS Infect Dis. 2020 May 8;6(5):1214-1227. doi: 10.1021/acsinfecdis.0c00015. Epub 2020 Mar 25. ACS Infect Dis. 2020. PMID: 32182432 Free PMC article.
-
Arginine Modulates Carbapenem Deactivation by OXA-24/40 in Acinetobacter baumannii.J Mol Biol. 2021 Sep 17;433(19):167150. doi: 10.1016/j.jmb.2021.167150. Epub 2021 Jul 14. J Mol Biol. 2021. PMID: 34271009 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
