

Melatonin inhibits nitric oxide production by microvascular endothelial cells in vivo and in vitro
- PMID: 17375079
- PMCID: PMC2013957
- DOI: 10.1038/sj.bjp.0707225
Melatonin inhibits nitric oxide production by microvascular endothelial cells in vivo and in vitro
Abstract
Background and purpose: We have previously shown that melatonin inhibits bradykinin-induced NO production by endothelial cells in vitro. The purpose of this investigation was to extend this observation to an in vivo condition and to explore the mechanism of action of melatonin.
Experimental approach: RT-PCR assays were performed with rat cultured endothelial cells. The putative effect of melatonin upon arteriolar tone was investigated by intravital microscopy while NO production by endothelial cells in vitro was assayed by fluorimetry, and intracellular Ca(2+) measurements were assayed by confocal microscopy.
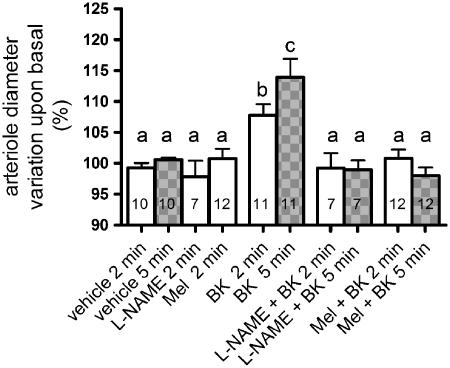
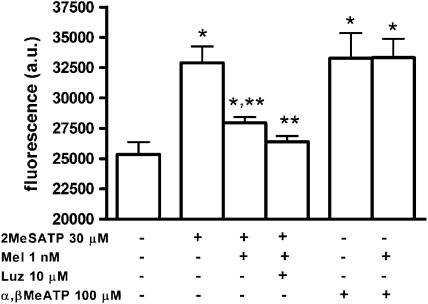
Key results: No expression of the mRNA for the melatonin synthesizing enzymes, arylalkylamine N-acetyltransferase and hydroxyindole-O-methyltransferase, or for the melatonin MT(2) receptor was detected in microvascular endothelial cells. Melatonin fully inhibited L-NAME-sensitive bradykinin-induced vasodilation and also inhibited NO production induced by histamine, carbachol and 2-methylthio ATP, but did not inhibit NO production induced by ATP or alpha, beta-methylene ATP. None of its inhibitory effects was prevented by the melatonin receptor antagonist, luzindole. In nominally Ca(2+)-free solution, melatonin reduced intracellular Ca(2+) mobilization induced by bradykinin (40%) and 2-methylthio ATP (62%) but not Ca(2+) mobilization induced by ATP.
Conclusions and implications: We have confirmed that melatonin inhibited NO production both in vivo and in vitro. In addition, the melatonin effect was selective for some G protein-coupled receptors and most probably reflects an inhibition of Ca(2+) mobilization from intracellular stores.
Figures




References
-
- Balcerczyk A, Soszynski M, Bartosz G. On the specificity of 4-amino-5-methylamino-2′,7′-difluorofluorescein as a probe for nitric oxide. Free Radic Biol Med. 2005;39:327–335. - PubMed
-
- Bauer PM, Fulton D, Boo YC, Sorescu GP, Kemp BE, Jo H, et al. Compensatory phosphorylation and protein–protein interactions revealed by loss of function and gain of function mutants of multiple serine phosphorylation sites in endothelial nitric-oxide synthase. J Biol Chem. 2003;278:14841–14849. - PubMed
-
- Benitez-King G, Huerto-Delgadillo L, Anton-Tay F. Binding of 3H-melatonin to calmodulin. Life Sci. 1993;53:201–207. - PubMed
-
- Beutelspacher SC, Tan PH, McClure MO, Larkin DF, Lechler RI, George AJ. Expression of indoleamine 2,3-dioxygenase (IDO) by endothelial cells: implications for the control of alloresponses. Am J Transplant. 2006;6:1320–1330. - PubMed
-
- Boutin JA, Audinot V, Ferry G, Delagrange P. Molecular tools to study melatonin pathways and actions. Trends Pharmacol Sci. 2005;26:412–419. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
