

Heteropolymeric triplex-based genomic assay to detect pathogens or single-nucleotide polymorphisms in human genomic samples
- PMID: 17375191
- PMCID: PMC1810429
- DOI: 10.1371/journal.pone.0000305
Heteropolymeric triplex-based genomic assay to detect pathogens or single-nucleotide polymorphisms in human genomic samples
Abstract
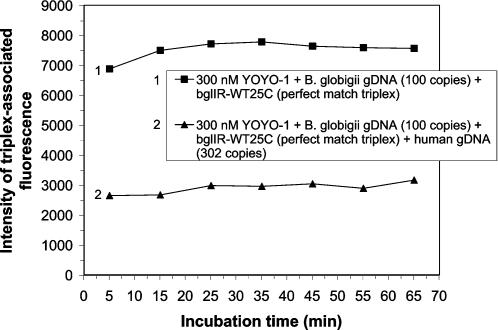
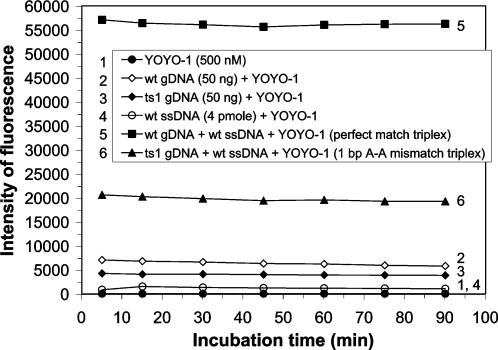
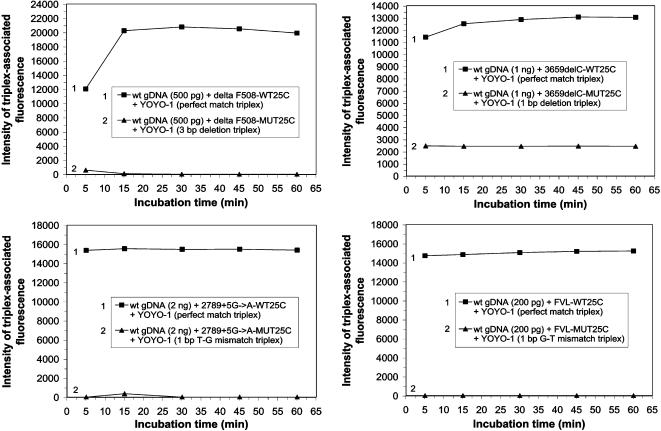
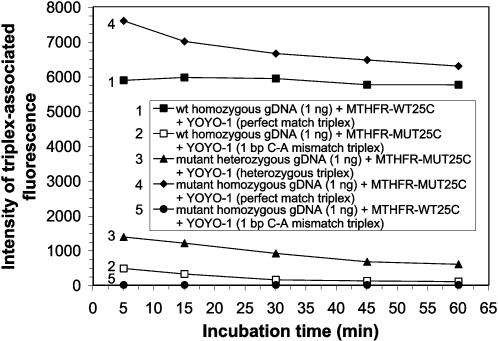
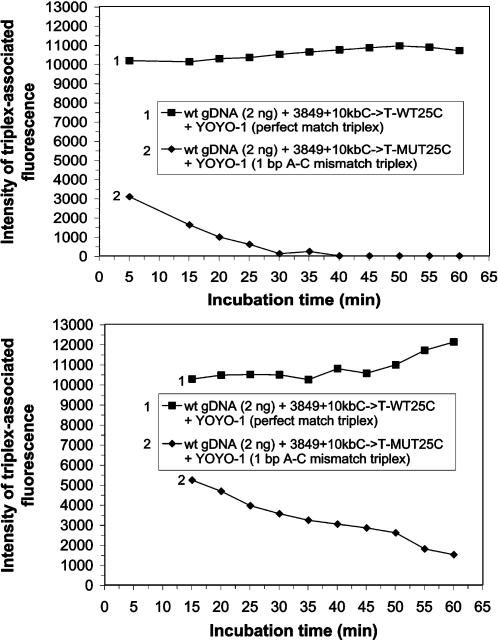
Human genomic samples are complex and are considered difficult to assay directly without denaturation or PCR amplification. We report the use of a base-specific heteropolymeric triplex, formed by native duplex genomic target and an oligonucleotide third strand probe, to assay for low copy pathogen genomes present in a sample also containing human genomic duplex DNA, or to assay human genomic duplex DNA for Single Nucleotide Polymorphisms (SNP), without PCR amplification. Wild-type and mutant probes are used to identify triplexes containing FVL G1691A, MTHFR C677T and CFTR mutations. The specific triplex structure forms rapidly at room temperature in solution and may be detected without a separation step. YOYO-1, a fluorescent bis-intercalator, promotes and signals the formation of the specific triplex. Genomic duplexes may be assayed homogeneously with single base pair resolution. The specific triple-stranded structures of the assay may approximate homologous recombination intermediates, which various models suggest may form in either the major or minor groove of the duplex. The bases of the stable duplex target are rendered specifically reactive to the bases of the probe because of the activity of intercalated YOYO-1, which is known to decondense duplex locally 1.3 fold. This may approximate the local decondensation effected by recombination proteins such as RecA in vivo. Our assay, while involving triplex formation, is sui generis, as it is not homopurine sequence-dependent, as are "canonical triplexes". Rather, the base pair-specific heteropolymeric triplex of the assay is conformation-dependent. The highly sensitive diagnostic assay we present allows for the direct detection of base sequence in genomic duplex samples, including those containing human genomic duplex DNA, thereby bypassing the inherent problems and cost associated with conventional PCR based diagnostic assays.
Conflict of interest statement
Figures
Similar articles
-
Evidence for a DNA triplex in a recombination-like motif: I. Recognition of Watson-Crick base pairs by natural bases in a high-stability triplex.J Mol Recognit. 2001 Mar-Apr;14(2):122-39. doi: 10.1002/jmr.528. J Mol Recognit. 2001. PMID: 11301482
-
Specific triplex binding capacity of mixed base sequence duplex nucleic acids used for single-nucleotide polymorphism detection.Genet Test. 2005 Summer;9(2):111-20. doi: 10.1089/gte.2005.9.111. Genet Test. 2005. PMID: 15943550
-
Promotion of duplex and triplex DNA formation by polycation comb-type copolymers.Methods Mol Med. 2001;65:209-24. doi: 10.1385/1-59259-139-6:209. Methods Mol Med. 2001. PMID: 21318757
-
The influence of intercalator binding on DNA triplex stability: correlation with effects on A-tract duplex structure.J Mol Recognit. 2004 Jul-Aug;17(4):277-85. doi: 10.1002/jmr.665. J Mol Recognit. 2004. PMID: 15227636 Review.
-
[Quantitative PCR in the diagnosis of Leishmania].Parassitologia. 2004 Jun;46(1-2):163-7. Parassitologia. 2004. PMID: 15305709 Review. Italian.
Cited by
-
Metagenomic DNA sequencing to quantify Mycobacterium tuberculosis DNA and diagnose tuberculosis.Sci Rep. 2022 Oct 10;12(1):16972. doi: 10.1038/s41598-022-21244-x. Sci Rep. 2022. PMID: 36216964 Free PMC article.
References
-
- Frank-Kamenetskii MD. DNA supercoiling and unusual structures. In: Cozzarelli NR, Wang JC, editors. DNA Topology and its Biological Effects. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 1990. pp. 185–215.
-
- Grigoriev M, Praseuth D, Robin P, Hemar A, Saison-Behmoaras T, et al. A triple helix-forming oligonucleotide-intercalator conjugate acts as a transcriptional repressor via inhibition of NF kappa B binding to interlelukin-2 receptor alpha-regulatory sequence. J Biol Chem. 1992;267:3389–3395. - PubMed
-
- Helene C, Thuong NT, Harel-Bellan A. Control of gene expression by triple helix-forming oligonucleotides. The antigene strategy. Ann NY Acad Sci. 1992;660:27–36. - PubMed
-
- Maher LJ, 3rd, Dervan PB, Wold B. Analysis of promoter-specific repression by triple-helical DNA complexes in a eukaryotic cell-free transcription system. Biochemistry. 1992;31:70–81. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
