

Progression from mitotic catastrophe to germ cell death in Caenorhabditis elegans lis-1 mutants requires the spindle checkpoint
- PMID: 17376425
- PMCID: PMC2000799
- DOI: 10.1016/j.ydbio.2007.02.024
Progression from mitotic catastrophe to germ cell death in Caenorhabditis elegans lis-1 mutants requires the spindle checkpoint
Abstract
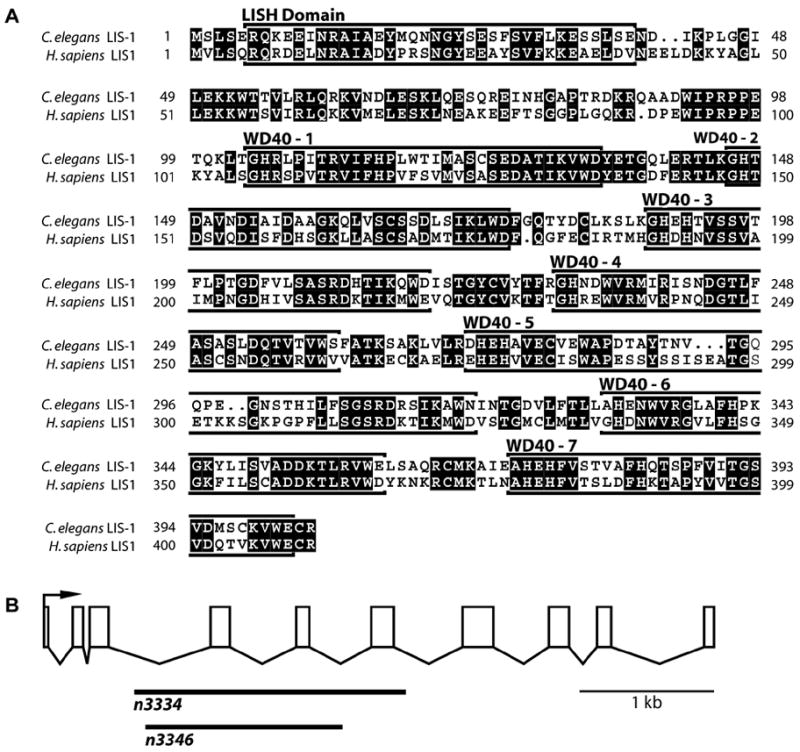
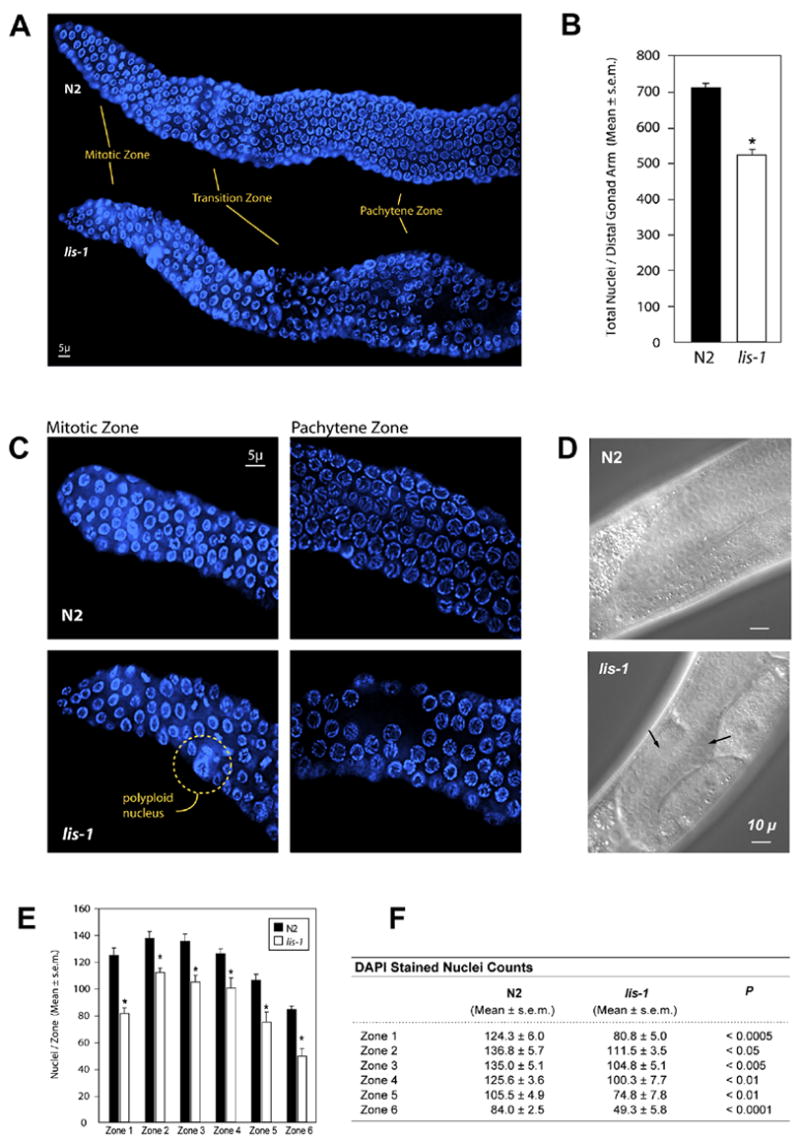
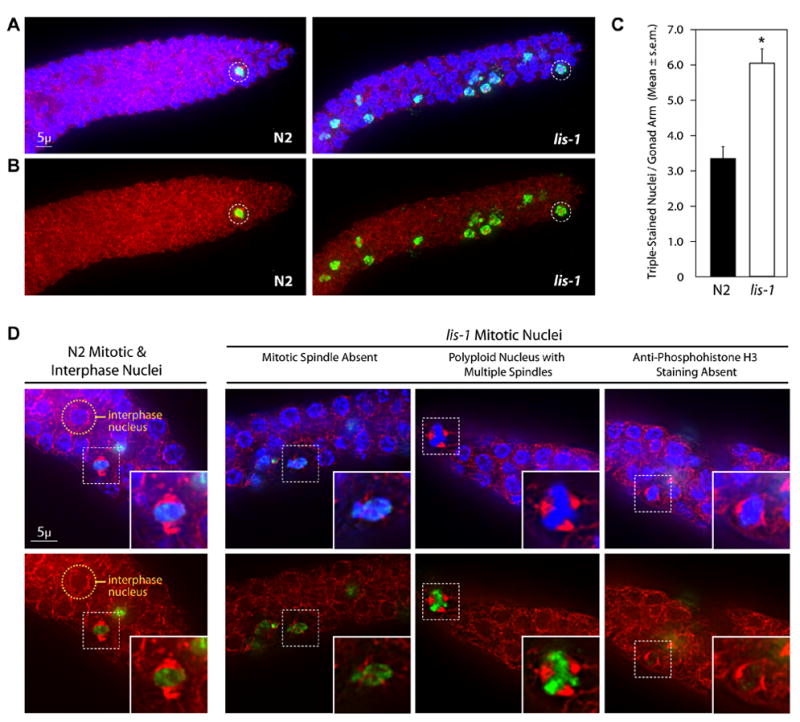
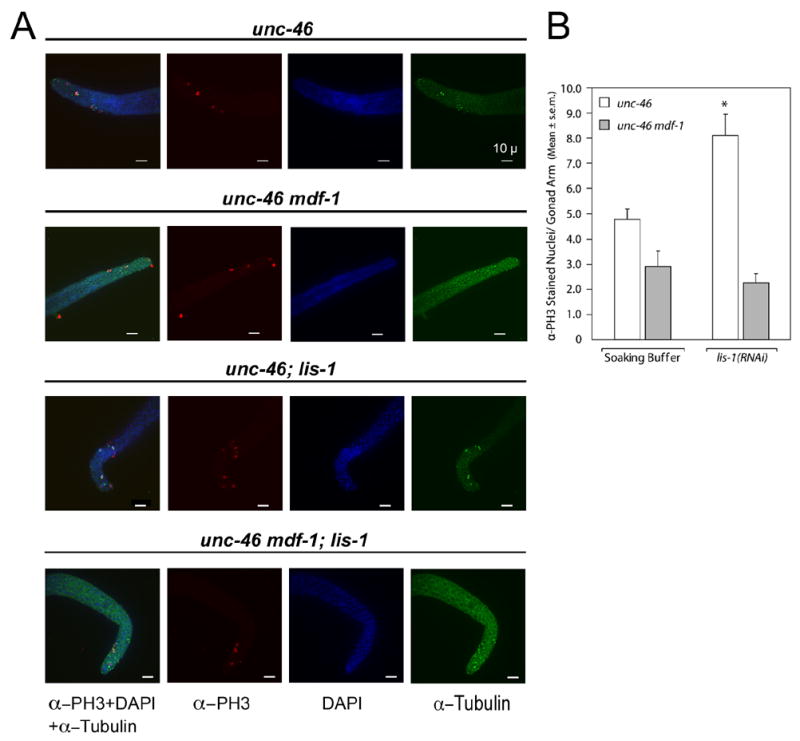
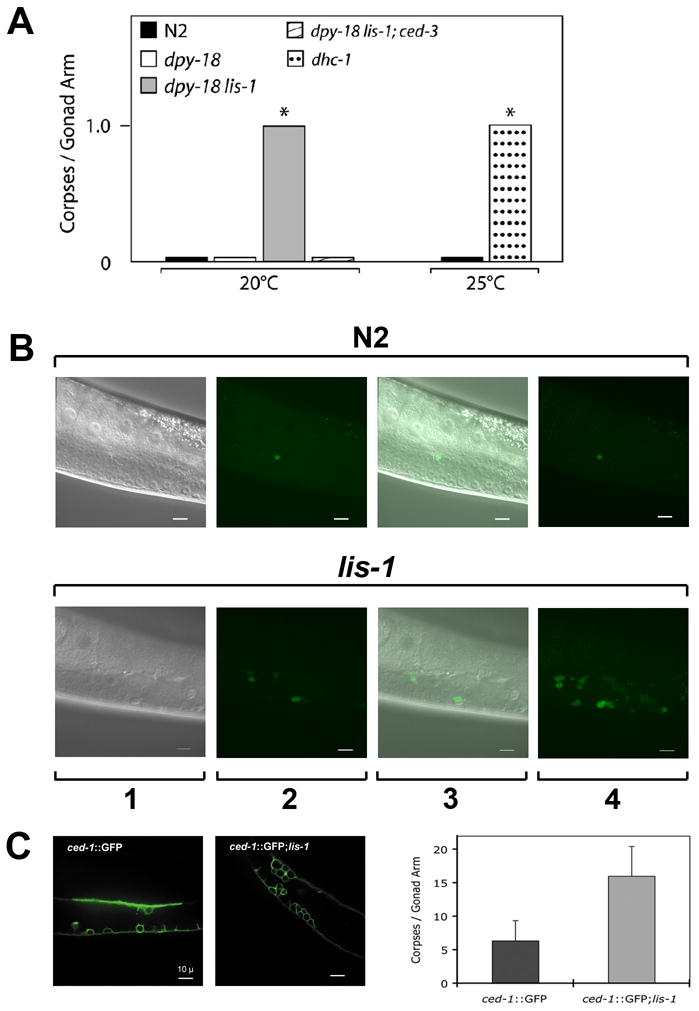
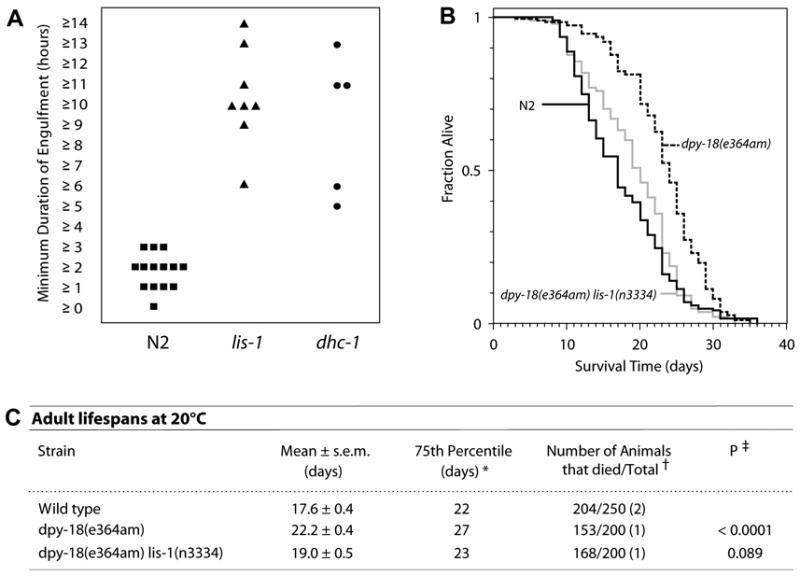
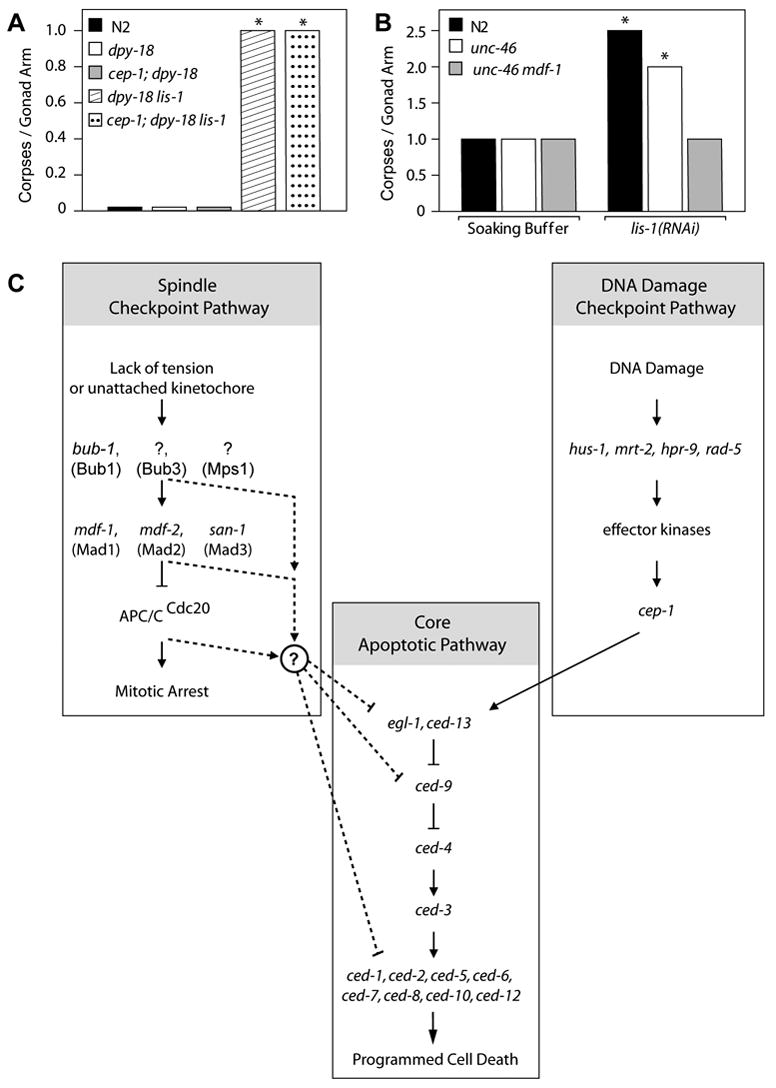
Deletion of the lissencephaly disease gene LIS-1 in humans causes an extreme disorganization of the brain associated with significant reduction in cortical neurons. Here we show that deletion or RNA interference (RNAi) of Caenorhabditis elegans lis-1 results in a reduction in germline nuclei and causes a variety of cellular, developmental, and neurological defects throughout development. Our analysis of the germline defects suggests that the reduction in nuclei number stems from dysfunctional mitotic spindles resulting in cell cycle arrest and eventually programmed cell death (apoptosis). Deletion of the spindle checkpoint gene mdf-1 blocks lis-1(lf)-induced cell cycle arrest and germline apoptosis, placing the spindle checkpoint pathway upstream of the programmed cell death pathway. These results suggest that apoptosis may contribute to the cell-sparse pathology of lissencephaly.
Figures
Similar articles
-
Suspended animation in C. elegans requires the spindle checkpoint.Science. 2003 Nov 7;302(5647):1038-41. doi: 10.1126/science.1089705. Science. 2003. PMID: 14605367
-
An essential function of the C. elegans ortholog of TPX2 is to localize activated aurora A kinase to mitotic spindles.Dev Cell. 2005 Aug;9(2):237-48. doi: 10.1016/j.devcel.2005.07.002. Dev Cell. 2005. PMID: 16054030
-
Suppressors of spindle checkpoint defect (such) mutants identify new mdf-1/MAD1 interactors in Caenorhabditis elegans.Genetics. 2007 Apr;175(4):1665-79. doi: 10.1534/genetics.106.067918. Epub 2007 Jan 21. Genetics. 2007. PMID: 17237515 Free PMC article.
-
Death and more: DNA damage response pathways in the nematode C. elegans.Cell Death Differ. 2004 Jan;11(1):21-8. doi: 10.1038/sj.cdd.4401340. Cell Death Differ. 2004. PMID: 14685168 Review.
-
[Research progress on spindle assembly checkpoint gene BubR1].Zhejiang Da Xue Xue Bao Yi Xue Ban. 2011 Jul;40(4):446-50. doi: 10.3785/j.issn.1008-9292.2011.04.018. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2011. PMID: 21845762 Review. Chinese.
Cited by
-
Key players in chromosome segregation in Caenorhabditis elegans.Front Biosci (Landmark Ed). 2009 Jan 1;14(4):1529-57. doi: 10.2741/3323. Front Biosci (Landmark Ed). 2009. PMID: 19273145 Free PMC article. Review.
-
Forward Genetic Screen in Caenorhabditis elegans Suggests F57A10.2 and acp-4 As Suppressors of C9ORF72 Related Phenotypes.Front Mol Neurosci. 2016 Nov 8;9:113. doi: 10.3389/fnmol.2016.00113. eCollection 2016. Front Mol Neurosci. 2016. PMID: 27877110 Free PMC article.
-
Regulators of the actin cytoskeleton mediate lethality in a Caenorhabditis elegans dhc-1 mutant.Mol Biol Cell. 2010 Aug 1;21(15):2707-20. doi: 10.1091/mbc.E09-07-0593. Epub 2010 Jun 16. Mol Biol Cell. 2010. PMID: 20554764 Free PMC article.
-
Pharmacogenetic analysis reveals a post-developmental role for Rac GTPases in Caenorhabditis elegans GABAergic neurotransmission.Genetics. 2009 Dec;183(4):1357-72. doi: 10.1534/genetics.109.106880. Epub 2009 Sep 21. Genetics. 2009. PMID: 19797046 Free PMC article.
-
Lis1 is essential for cortical microtubule organization and desmosome stability in the epidermis.J Cell Biol. 2011 Aug 22;194(4):631-42. doi: 10.1083/jcb.201104009. Epub 2011 Aug 15. J Cell Biol. 2011. PMID: 21844209 Free PMC article.
References
-
- Bhalla KN. Microtubule-targeted anticancer agents and apoptosis. Oncogene. 2003;22:9075–9086. - PubMed
-
- Cockell MM, Baumer K, Gonczy P. lis-1 is required for dynein-dependent cell division processes in C. elegans embryos. J Cell Sci. 2004;117:4571–4582. - PubMed
-
- Davidson FF, Steller H. Blocking apoptosis prevents blindness in Drosophila retinal degeneration mutants. Nature. 1998;391:587–591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
