

Function of the ATR N-terminal domain revealed by an ATM/ATR chimera
- PMID: 17376433
- PMCID: PMC1855264
- DOI: 10.1016/j.yexcr.2007.02.015
Function of the ATR N-terminal domain revealed by an ATM/ATR chimera
Abstract
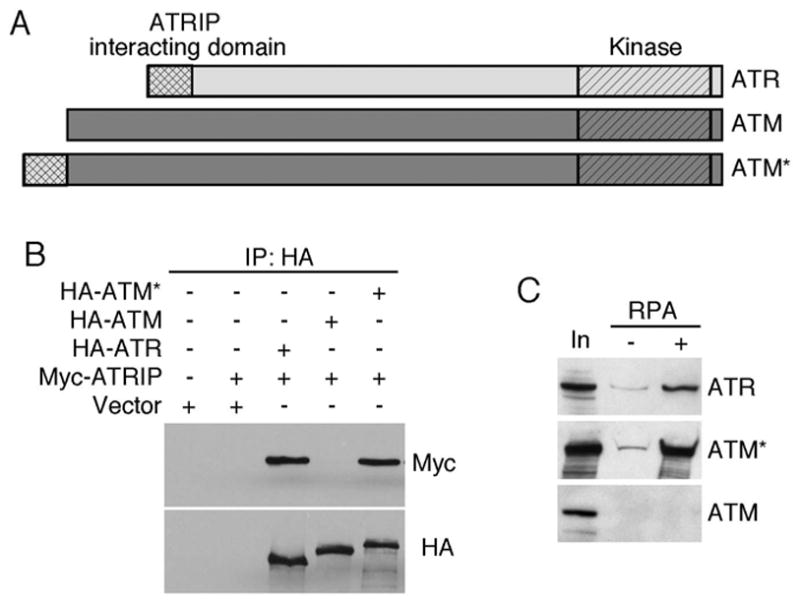
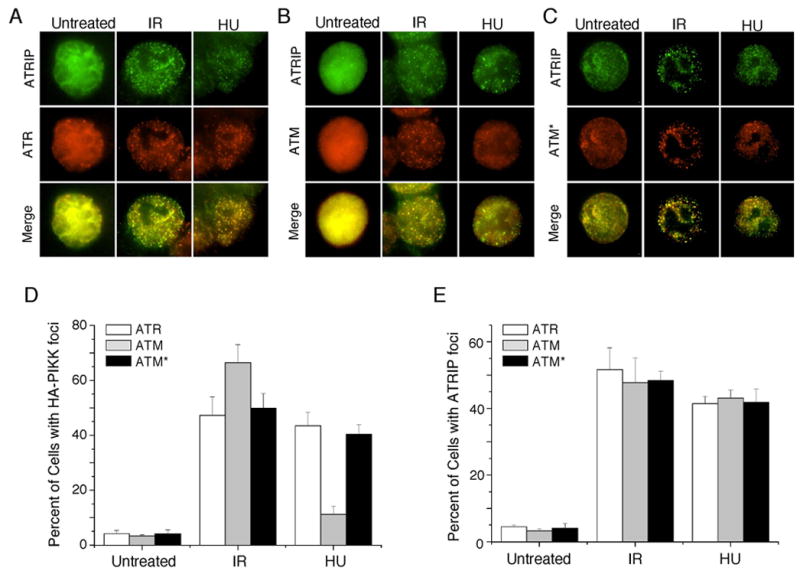
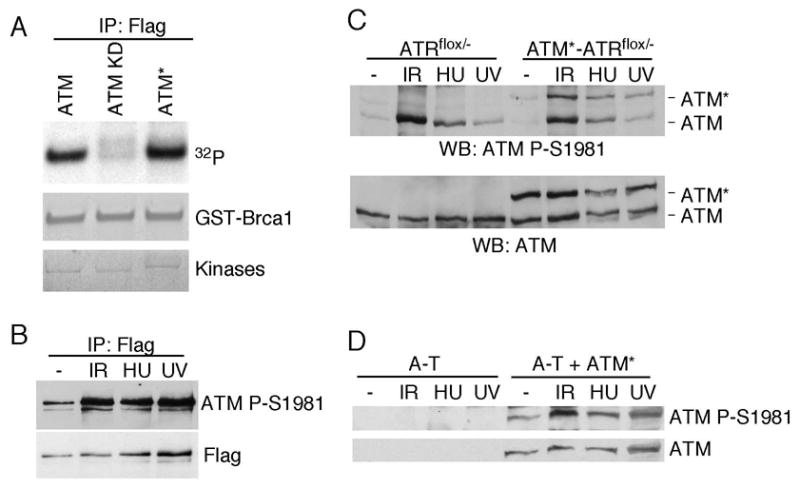
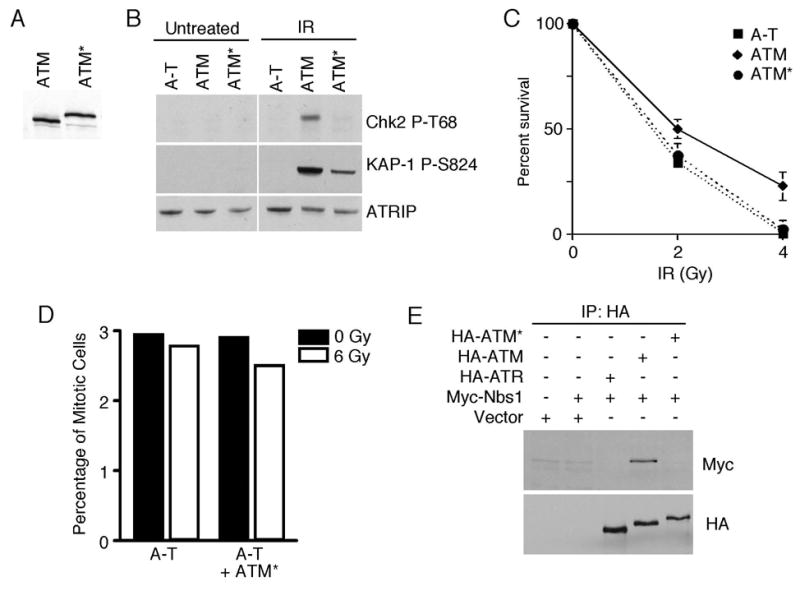
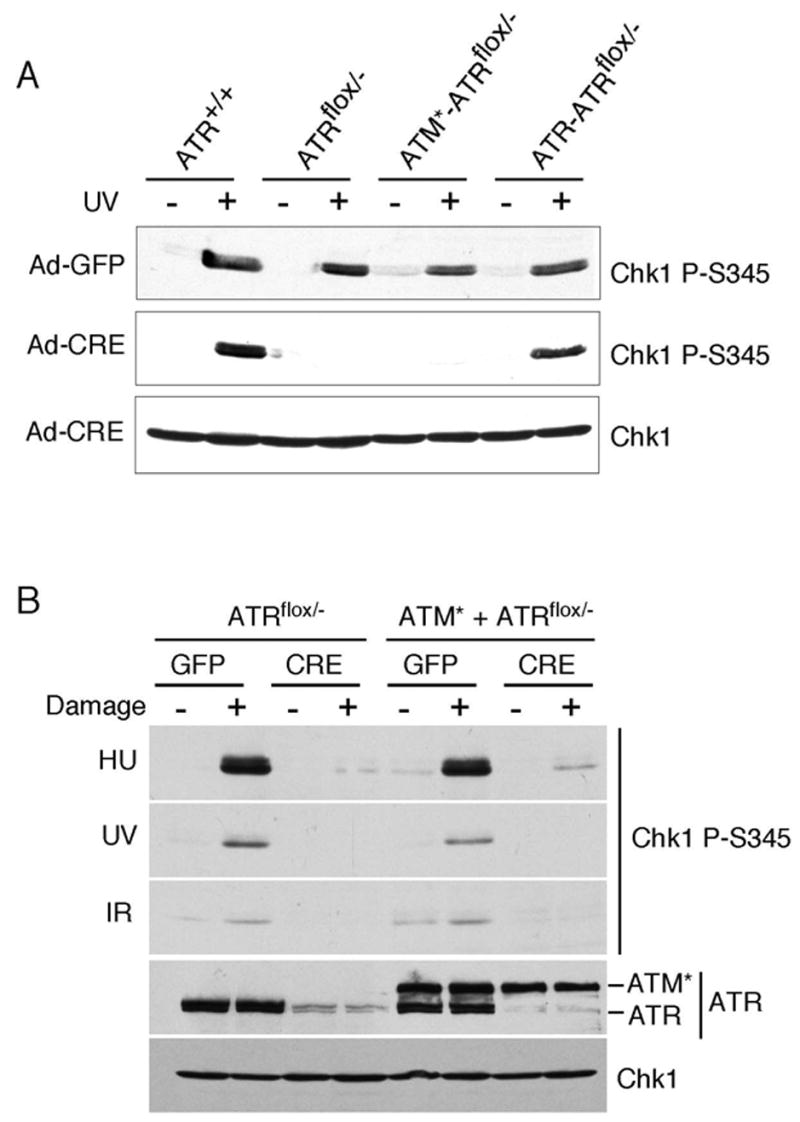
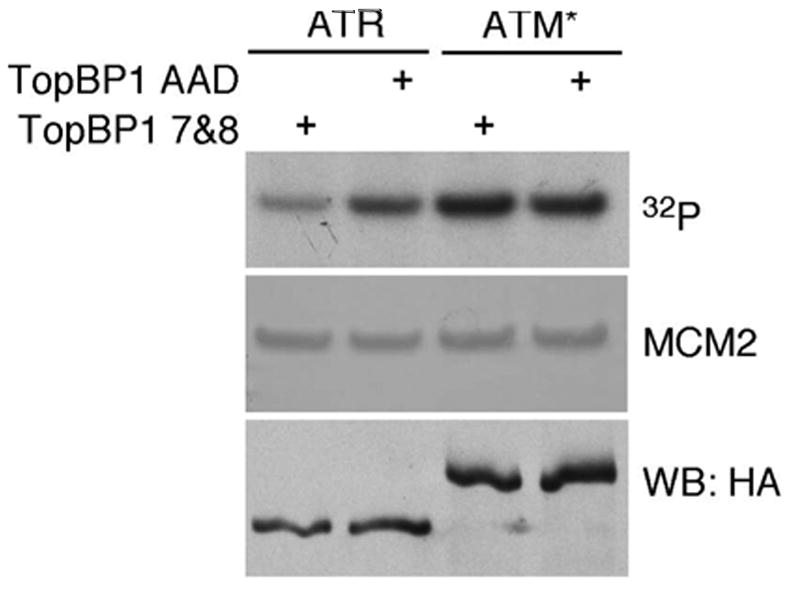
The ATM and ATR kinases function at the apex of checkpoint signaling pathways. These kinases share significant sequence similarity, phosphorylate many of the same substrates, and have overlapping roles in initiating cell cycle checkpoints. However, they sense DNA damage through distinct mechanisms. ATR primarily senses single stranded DNA (ssDNA) through its interaction with ATRIP, and ATM senses double strand breaks through its interaction with Nbs1. We determined that the N-terminus of ATR contains a domain that binds ATRIP. Attaching this domain to ATM allowed the fusion protein (ATM*) to bind ATRIP and associate with RPA-coated ssDNA. ATM* also gained the ability to localize efficiently to stalled replication forks as well as double strand breaks. Despite having normal kinase activity when tested in vitro and being phosphorylated on S1981 in vivo, ATM* is defective in checkpoint signaling and does not complement cellular deficiencies in either ATM or ATR. These data indicate that the N-terminus of ATR is sufficient to bind ATRIP and to promote localization to sites of replication stress.
Figures
Similar articles
-
ATRIP binding to replication protein A-single-stranded DNA promotes ATR-ATRIP localization but is dispensable for Chk1 phosphorylation.Mol Biol Cell. 2005 May;16(5):2372-81. doi: 10.1091/mbc.e04-11-1006. Epub 2005 Mar 2. Mol Biol Cell. 2005. PMID: 15743907 Free PMC article.
-
Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes.Science. 2003 Jun 6;300(5625):1542-8. doi: 10.1126/science.1083430. Science. 2003. PMID: 12791985
-
Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage.Nature. 2005 Mar 31;434(7033):605-11. doi: 10.1038/nature03442. Epub 2005 Mar 2. Nature. 2005. PMID: 15758953
-
The ATM-dependent DNA damage signaling pathway.Cold Spring Harb Symp Quant Biol. 2005;70:99-109. doi: 10.1101/sqb.2005.70.002. Cold Spring Harb Symp Quant Biol. 2005. PMID: 16869743 Review.
-
Activation of ATR-related protein kinase upon DNA damage recognition.Curr Genet. 2020 Apr;66(2):327-333. doi: 10.1007/s00294-019-01039-w. Epub 2019 Oct 17. Curr Genet. 2020. PMID: 31624858 Free PMC article. Review.
Cited by
-
A TRilogy of ATR's Non-Canonical Roles Throughout the Cell Cycle and Its Relation to Cancer.Cancers (Basel). 2024 Oct 19;16(20):3536. doi: 10.3390/cancers16203536. Cancers (Basel). 2024. PMID: 39456630 Free PMC article. Review.
-
Mutation of serine 1333 in the ATR HEAT repeats creates a hyperactive kinase.PLoS One. 2014 Jun 5;9(6):e99397. doi: 10.1371/journal.pone.0099397. eCollection 2014. PLoS One. 2014. PMID: 24901225 Free PMC article.
-
TopBP1 activates ATR through ATRIP and a PIKK regulatory domain.Genes Dev. 2008 Jun 1;22(11):1478-89. doi: 10.1101/gad.1666208. Genes Dev. 2008. PMID: 18519640 Free PMC article.
-
ATR: an essential regulator of genome integrity.Nat Rev Mol Cell Biol. 2008 Aug;9(8):616-27. doi: 10.1038/nrm2450. Epub 2008 Jul 2. Nat Rev Mol Cell Biol. 2008. PMID: 18594563 Free PMC article. Review.
-
The role of MRN in the S-phase DNA damage checkpoint is independent of its Ctp1-dependent roles in double-strand break repair and checkpoint signaling.Mol Biol Cell. 2009 Apr;20(7):2096-107. doi: 10.1091/mbc.e08-09-0986. Epub 2009 Feb 11. Mol Biol Cell. 2009. PMID: 19211838 Free PMC article.
References
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, Orntoft T, Lukas J, Bartek J. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
-
- Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, Vassiliou LV, Kolettas E, Niforou K, Zoumpourlis VC, Takaoka M, Nakagawa H, Tort F, Fugger K, Johansson F, Sehested M, Andersen CL, Dyrskjot L, Orntoft T, Lukas J, Kittas C, Helleday T, Halazonetis TD, Bartek J, Gorgoulis VG. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature. 2006;444:633–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
