

Conformation-specific antibodies targeting the trimer-of-hairpins motif of the human T-cell leukemia virus type 1 transmembrane glycoprotein recognize the viral envelope but fail to neutralize viral entry
- PMID: 17376912
- PMCID: PMC1900303
- DOI: 10.1128/JVI.02544-06
Conformation-specific antibodies targeting the trimer-of-hairpins motif of the human T-cell leukemia virus type 1 transmembrane glycoprotein recognize the viral envelope but fail to neutralize viral entry
Abstract
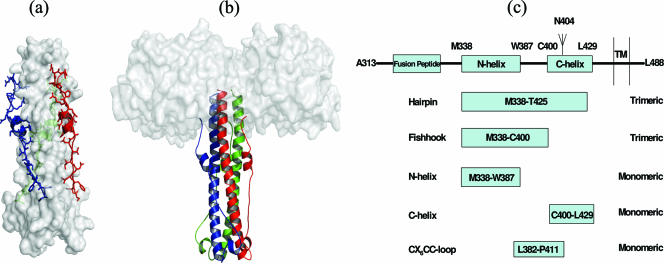
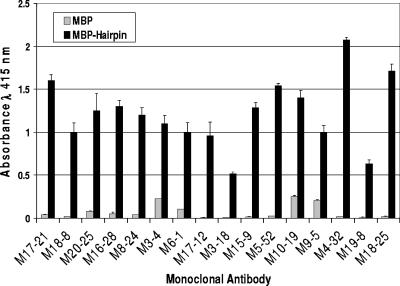
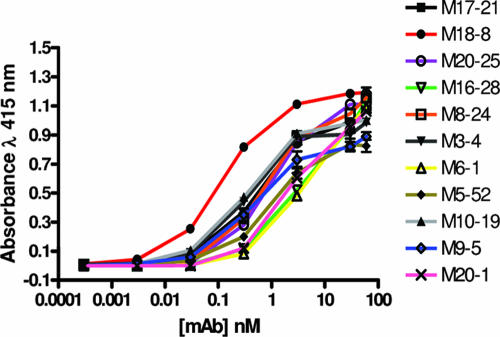
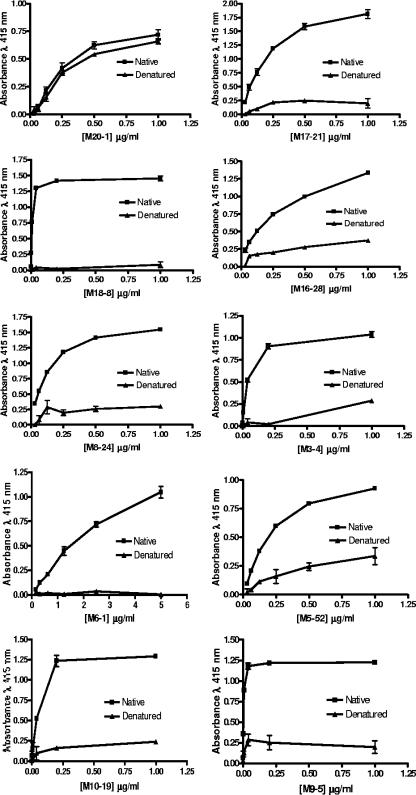
Human T-cell leukemia virus type 1 (HTLV-1) entry into cells is dependent upon the viral envelope glycoprotein-catalyzed fusion of the viral and cellular membranes. Following receptor activation of the envelope, the transmembrane glycoprotein (TM) is thought to undergo a series of fusogenic conformational transitions through a rod-like prehairpin intermediate to a compact trimer-of-hairpins structure. Importantly, synthetic peptides that interfere with the conformational changes of TM are potent inhibitors of membrane fusion and HTLV-1 entry, suggesting that TM is a valid target for antiviral therapy. To assess the utility of TM as a vaccine target and to explore further the function of TM in HTLV-1 pathogenesis, we have begun to examine the immunological properties of TM. Here we demonstrate that a recombinant trimer-of-hairpins form of the TM ectodomain is strongly immunogenic. Monoclonal antibodies raised against the TM immunogen specifically bind to trimeric forms of TM, including structures thought to be important for membrane fusion. Importantly, these antibodies recognize the envelope on virally infected cells but, surprisingly, fail to neutralize envelope-mediated membrane fusion or infection by pseudotyped viral particles. Our data imply that, even in the absence of overt membrane fusion, there are multiple forms of TM on virally infected cells and that some of these display fusion-associated structures. Finally, we demonstrate that many of the antibodies possess the ability to recruit complement to TM, suggesting that envelope-derived immunogens capable of eliciting a combination of neutralizing and complement-fixing antibodies would be of value as subunit vaccines for intervention in HTLV infections.
Figures
Similar articles
-
An antibody that blocks human T-cell leukemia virus type 1 six-helix-bundle formation in vitro identified by a novel assay for inhibitors of envelope function.J Gen Virol. 2007 Feb;88(Pt 2):660-669. doi: 10.1099/vir.0.82390-0. J Gen Virol. 2007. PMID: 17251585
-
Antibodies to the envelope glycoprotein of human T cell leukemia virus type 1 robustly activate cell-mediated cytotoxic responses and directly neutralize viral infectivity at multiple steps of the entry process.J Immunol. 2011 Jul 1;187(1):361-71. doi: 10.4049/jimmunol.1100070. Epub 2011 Jun 6. J Immunol. 2011. PMID: 21646298
-
Resistance to neutralization by antibodies targeting the coiled coil of fusion-active envelope is a common feature of retroviruses.J Biol Chem. 2007 Dec 14;282(50):36724-35. doi: 10.1074/jbc.M706827200. Epub 2007 Oct 16. J Biol Chem. 2007. PMID: 17940280
-
The HTLV-I envelope glycoproteins: structure and functions.J Acquir Immune Defic Syndr Hum Retrovirol. 1996;13 Suppl 1:S85-91. doi: 10.1097/00042560-199600001-00015. J Acquir Immune Defic Syndr Hum Retrovirol. 1996. PMID: 8797709 Review.
-
[Chemokine receptors and its importance in the replication cycle of human immunodeficiency virus: clinical and therapeutic implications].Acta Med Port. 2008 Sep-Oct;21(5):497-504. Epub 2009 Jan 16. Acta Med Port. 2008. PMID: 19187693 Review. Portuguese.
Cited by
-
Cellular Factors Involved in HTLV-1 Entry and Pathogenicit.Front Microbiol. 2012 Jun 21;3:222. doi: 10.3389/fmicb.2012.00222. eCollection 2012. Front Microbiol. 2012. PMID: 22737146 Free PMC article.
-
Advances in preventive vaccine development against HTLV-1 infection: A systematic review of the last 35 years.Front Immunol. 2023 Feb 13;14:1073779. doi: 10.3389/fimmu.2023.1073779. eCollection 2023. Front Immunol. 2023. PMID: 36860854 Free PMC article.
-
Molecular aspects of HTLV-1 entry: functional domains of the HTLV-1 surface subunit (SU) and their relationships to the entry receptors.Viruses. 2011 Jun;3(6):794-810. doi: 10.3390/v3060794. Epub 2011 Jun 15. Viruses. 2011. PMID: 21994754 Free PMC article. Review.
-
Highly specific inhibition of leukaemia virus membrane fusion by interaction of peptide antagonists with a conserved region of the coiled coil of envelope.Retrovirology. 2008 Aug 4;5:70. doi: 10.1186/1742-4690-5-70. Retrovirology. 2008. PMID: 18680566 Free PMC article.
References
-
- Arvin, A. M., and H. B. Greenberg. 2006. New viral vaccines. Virology 344:240-249. - PubMed
-
- Baba, E., M. Nakamura, Y. Tanaka, M. Kuroki, Y. Itoyama, S. Nakano, and Y. Niho. 1993. Multiple neutralizing B-cell epitopes of human T-cell leukemia virus type 1 (HTLV-1) identified by human monoclonal antibodies. A basis for the design of an HTLV-1 peptide vaccine. J. Immunol. 151:1013-1024. - PubMed
-
- Borrow, P., H. Lewicki, X. Wei, M. S. Horwitz, N. Peffer, H. Meyers, J. A. Nelson, J. E. Gairin, B. H. Hahn, M. B. Oldstone, and G. M. Shaw. 1997. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med. 3:205-211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
