

Lhx6 activity is required for the normal migration and specification of cortical interneuron subtypes
- PMID: 17376969
- PMCID: PMC6672459
- DOI: 10.1523/JNEUROSCI.3055-06.2007
Lhx6 activity is required for the normal migration and specification of cortical interneuron subtypes
Abstract
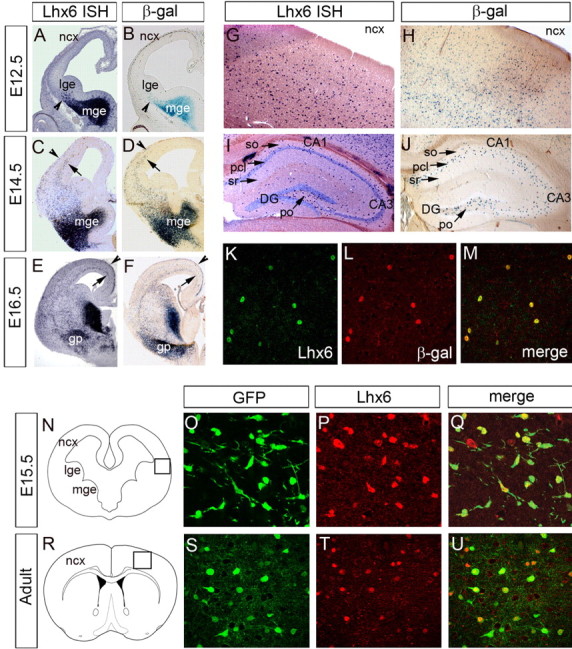
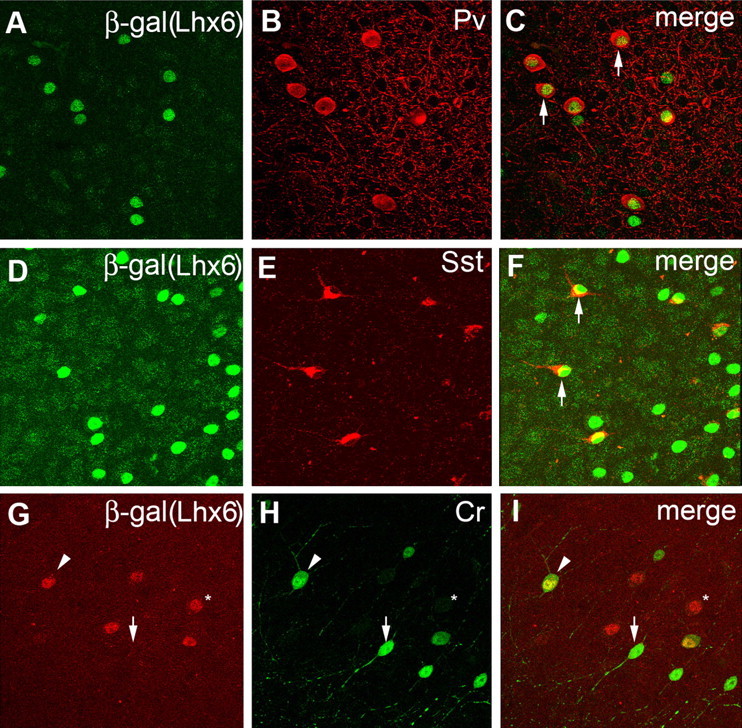
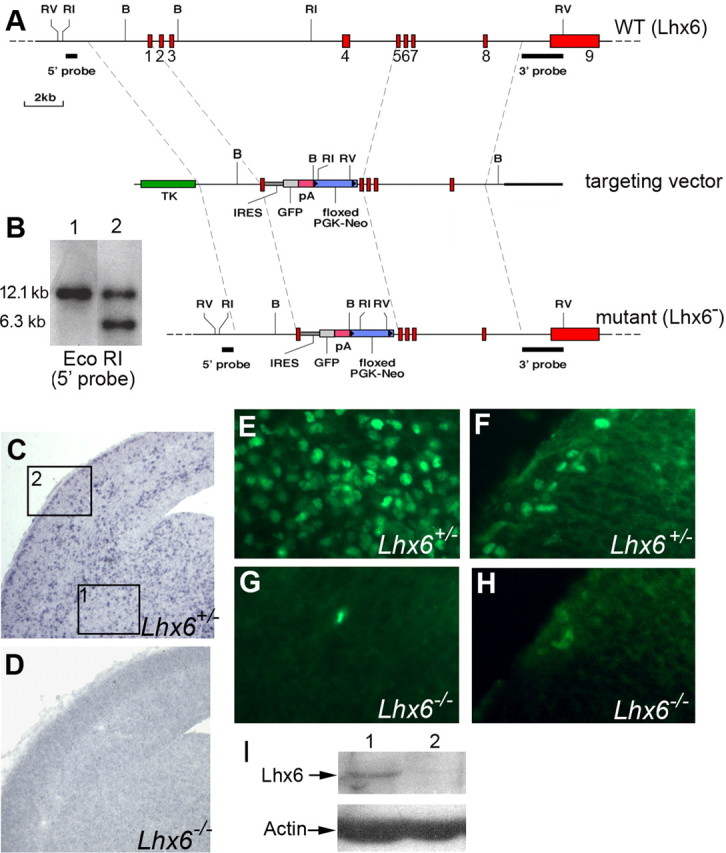
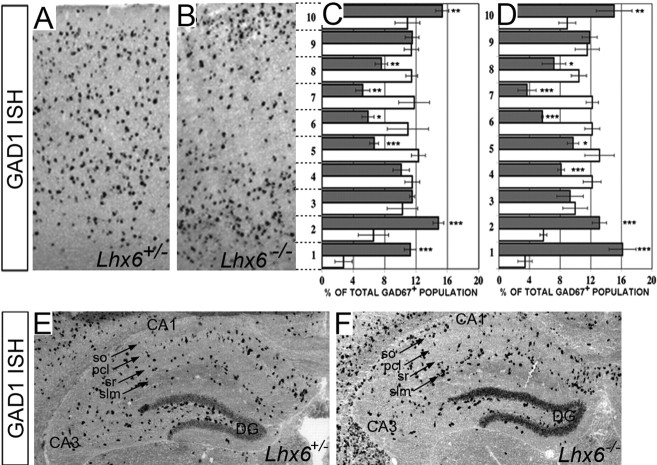
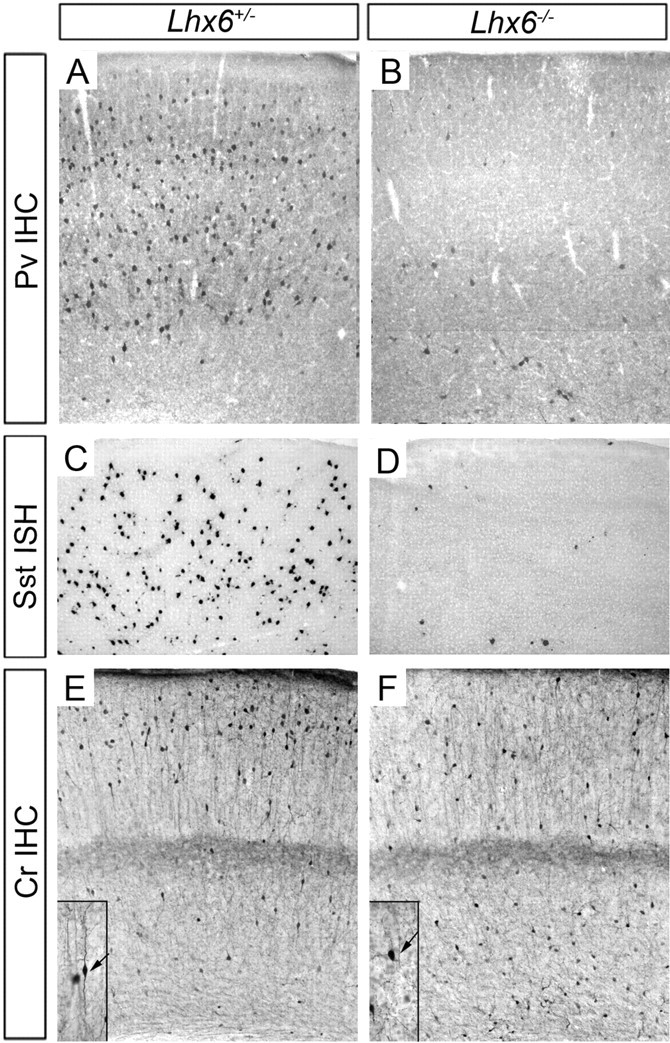
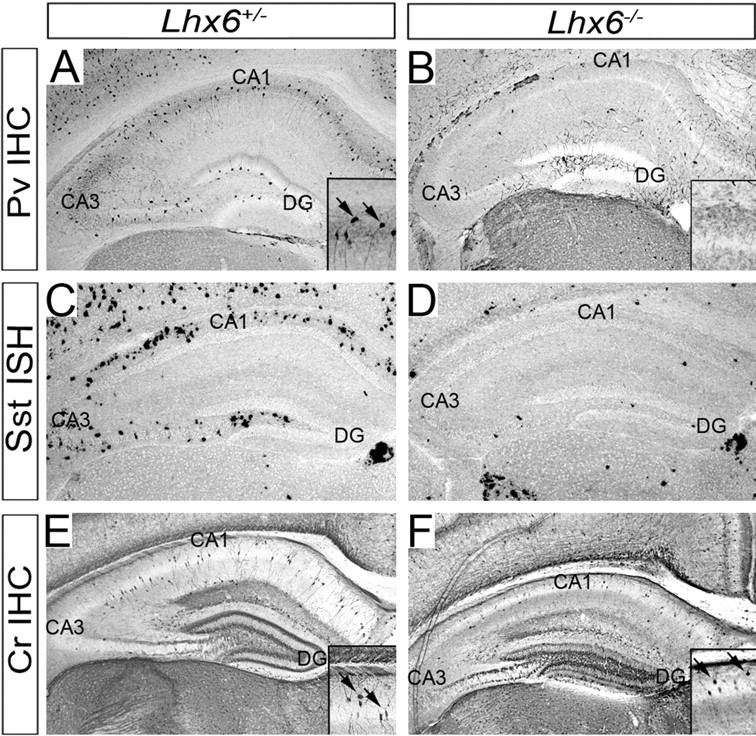
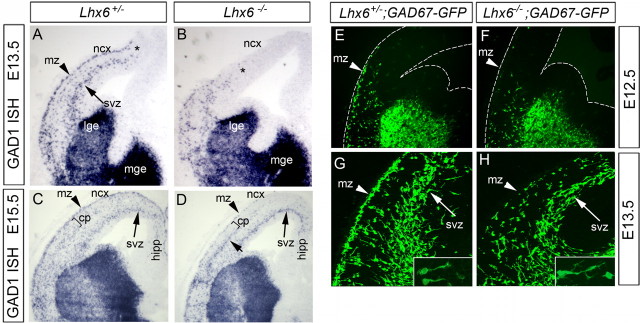
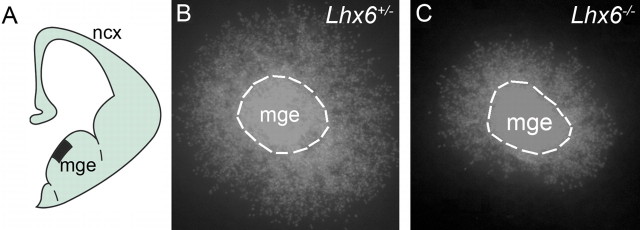
The cerebral cortex contains two main neuronal cell populations, the excitatory glutamatergic (pyramidal) neurons and the inhibitory interneurons, which synthesize GABA and constitute 20-30% of all cortical neurons. In contrast to the mostly homogeneous population of projection neurons, cortical interneurons are characterized by remarkable morphological, molecular, and functional diversity. Among the markers that have been used to classify cortical interneurons are the calcium-binding proteins parvalbumin and calretinin and the neuropeptide somatostatin, which in rodents identify mostly nonoverlapping interneuron subpopulations. Pyramidal neurons are born during embryogenesis in the ventricular zone of the dorsal telencephalon, whereas cortical interneurons are generated in the subpallium and reach the cortex by tangential migration. On completion of tangential migration, cortical interneurons switch to a radial mode of migration and enter the cortical plate. Although the mechanisms that control the generation of interneuron diversity are currently unknown, it has been proposed that their site of origin in the ventral forebrain determines their specification into defined neurochemical subgroups. Here, we show that Lhx6, a gene induced in the medial ganglionic eminence and maintained in parvalbumin- and somatostatin-positive interneurons, is required for the specification of these neuronal subtypes in the neocortex and the hippocampus. We also show that Lhx6 activity is required for the normal tangential and radial migration of GABAergic interneurons in the cortex.
Figures
Similar articles
-
Origins of cortical interneuron subtypes.J Neurosci. 2004 Mar 17;24(11):2612-22. doi: 10.1523/JNEUROSCI.5667-03.2004. J Neurosci. 2004. PMID: 15028753 Free PMC article.
-
Distinct molecular pathways for development of telencephalic interneuron subtypes revealed through analysis of Lhx6 mutants.J Comp Neurol. 2008 Sep 1;510(1):79-99. doi: 10.1002/cne.21772. J Comp Neurol. 2008. PMID: 18613121 Free PMC article.
-
The LIM homeodomain protein Lhx6 regulates maturation of interneurons and network excitability in the mammalian cortex.Cereb Cortex. 2013 Aug;23(8):1811-23. doi: 10.1093/cercor/bhs159. Epub 2012 Jun 17. Cereb Cortex. 2013. PMID: 22710612 Free PMC article.
-
The development of MGE-derived cortical interneurons: An Lhx6 tale.Int J Dev Biol. 2022;66(1-2-3):43-49. doi: 10.1387/ijdb.210185md. Int J Dev Biol. 2022. PMID: 34881792 Review.
-
Cortical interneuron fate determination: diverse sources for distinct subtypes?Cereb Cortex. 2003 Jun;13(6):670-6. doi: 10.1093/cercor/13.6.670. Cereb Cortex. 2003. PMID: 12764043 Review.
Cited by
-
Interneuron Origins in the Embryonic Porcine Medial Ganglionic Eminence.J Neurosci. 2021 Apr 7;41(14):3105-3119. doi: 10.1523/JNEUROSCI.2738-20.2021. Epub 2021 Feb 26. J Neurosci. 2021. PMID: 33637558 Free PMC article.
-
Cortical GABA markers identify a molecular subtype of psychotic and bipolar disorders.Psychol Med. 2016 Sep;46(12):2501-12. doi: 10.1017/S0033291716001446. Epub 2016 Jun 22. Psychol Med. 2016. PMID: 27328999 Free PMC article.
-
Association of Genetic Variation in the 3'UTR of LHX6, IMMP2L, and AADAC With Tourette Syndrome.Front Neurol. 2020 Aug 14;11:803. doi: 10.3389/fneur.2020.00803. eCollection 2020. Front Neurol. 2020. PMID: 32922348 Free PMC article.
-
Use of "MGE enhancers" for labeling and selection of embryonic stem cell-derived medial ganglionic eminence (MGE) progenitors and neurons.PLoS One. 2013 May 1;8(5):e61956. doi: 10.1371/journal.pone.0061956. Print 2013. PLoS One. 2013. PMID: 23658702 Free PMC article.
-
Interneuron Progenitor Transplantation to Treat CNS Dysfunction.Front Neural Circuits. 2016 Aug 17;10:64. doi: 10.3389/fncir.2016.00064. eCollection 2016. Front Neural Circuits. 2016. PMID: 27582692 Free PMC article. Review.
References
-
- Alcantara S, Soriano E, Ferrer I. Thalamic and basal forebrain afferents modulate the development of parvalbumin and calbindin D28k immunoreactivity in the barrel cortex of the rat. Eur J Neurosci. 1996;8:1522–1534. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Anderson SA, Marin O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases