

Segment specificity of load signal processing depends on walking direction in the stick insect leg muscle control system
- PMID: 17376989
- PMCID: PMC6672458
- DOI: 10.1523/JNEUROSCI.5202-06.2007
Segment specificity of load signal processing depends on walking direction in the stick insect leg muscle control system
Abstract
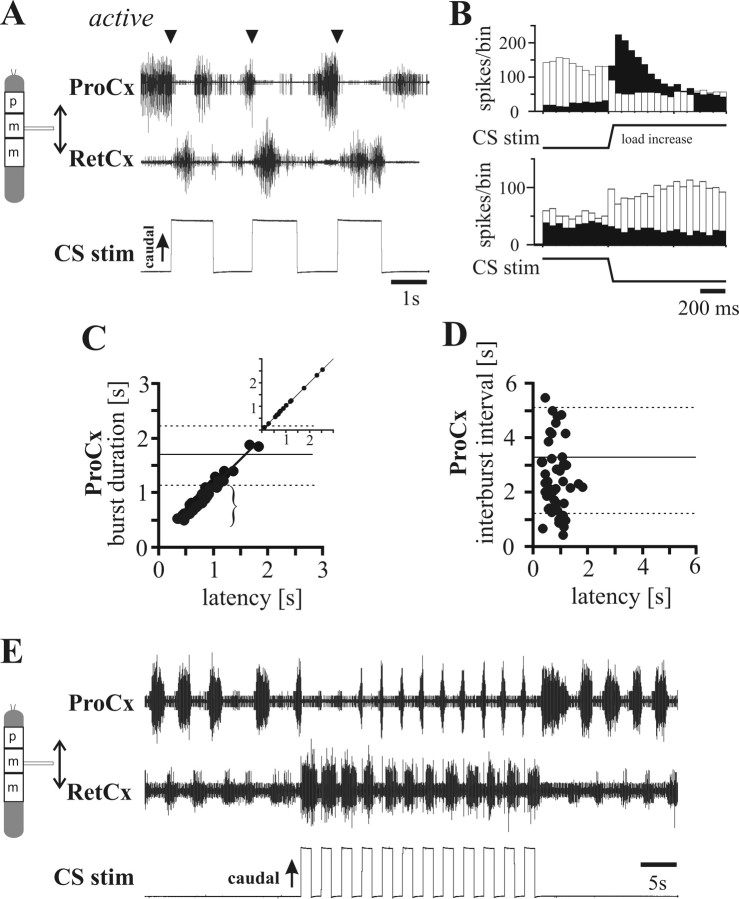
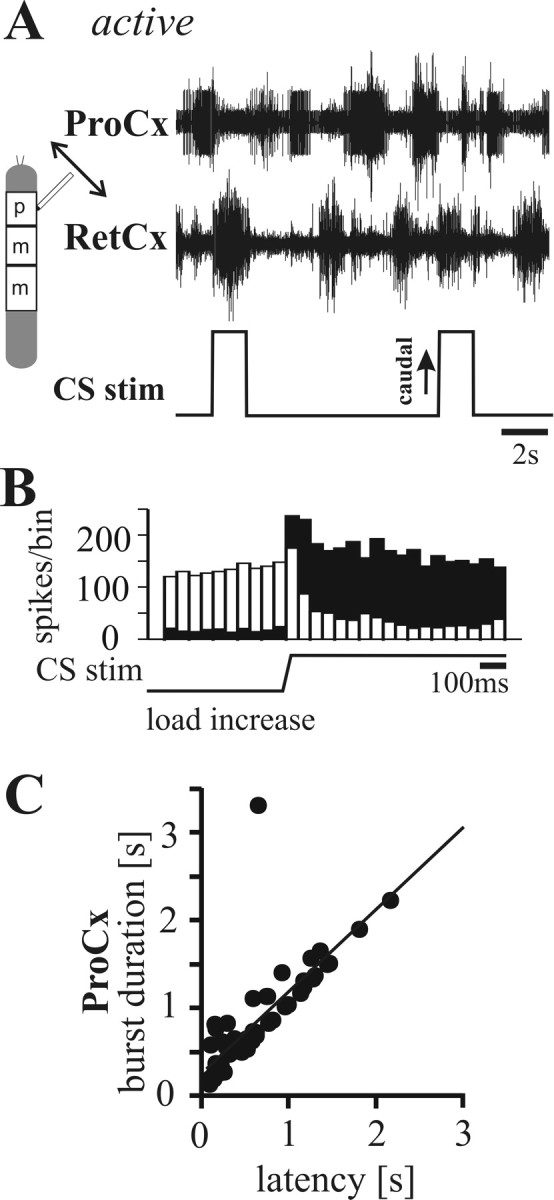
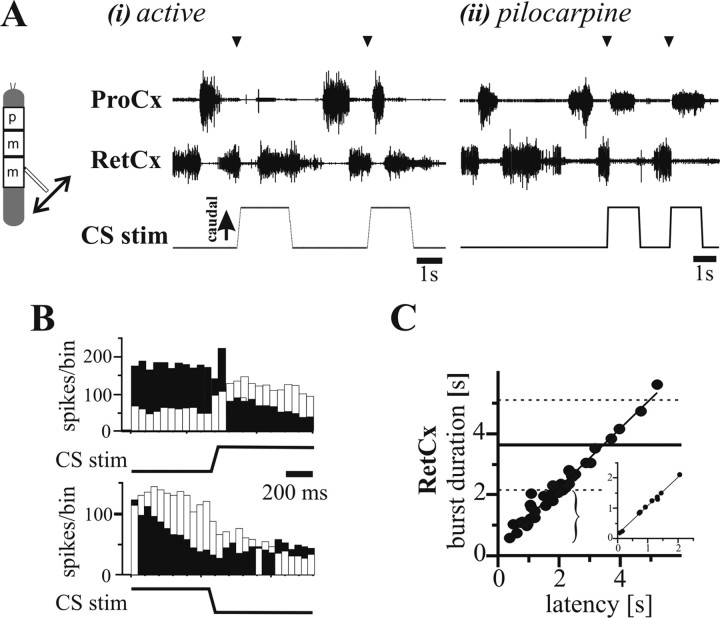
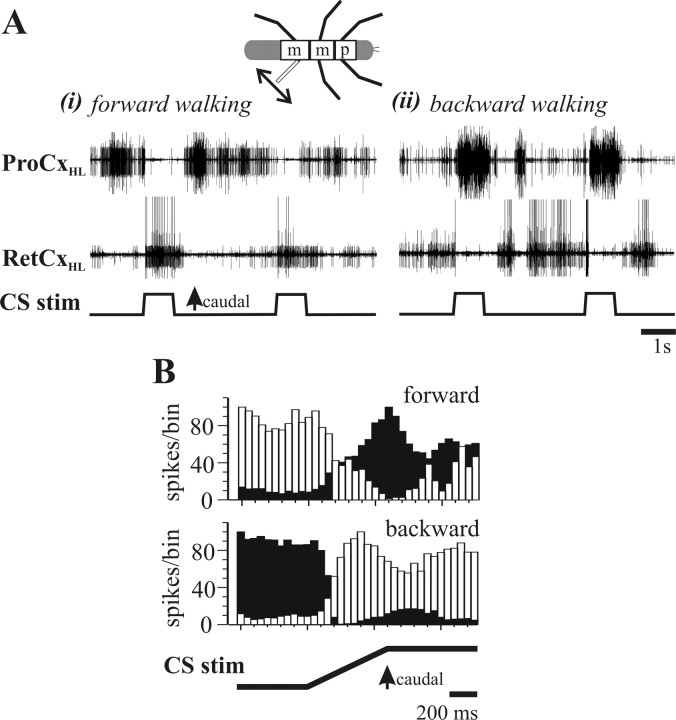
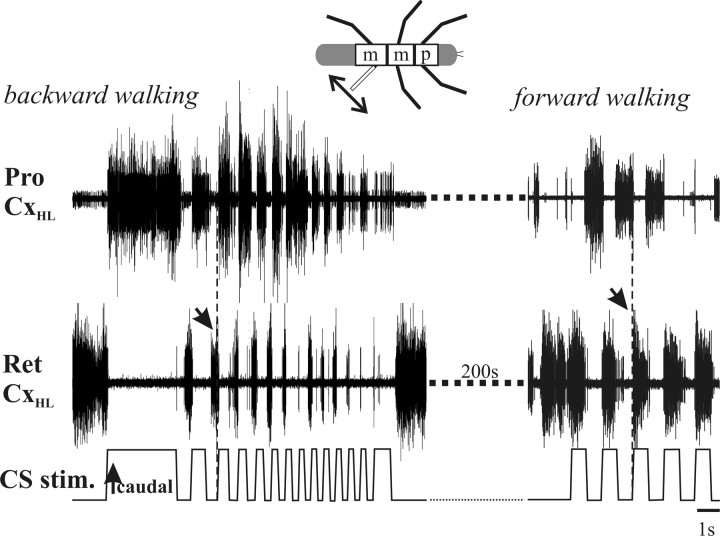
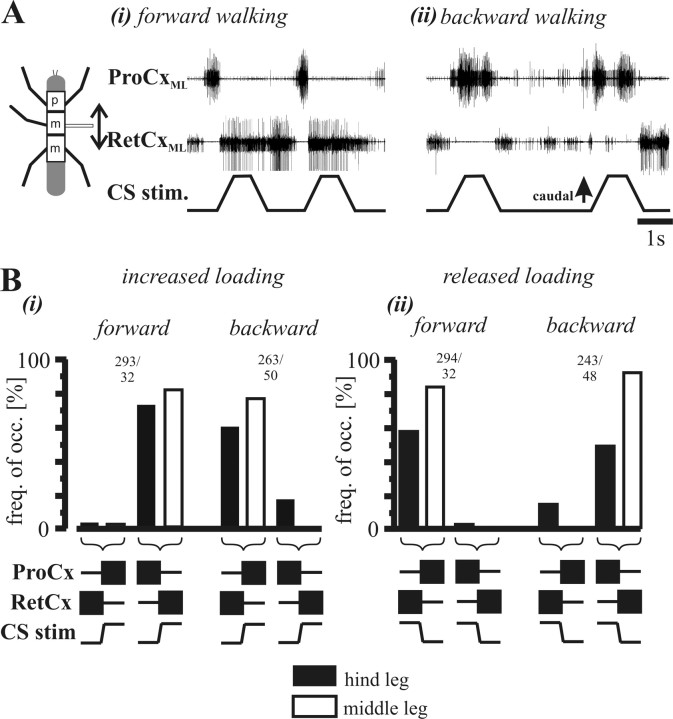
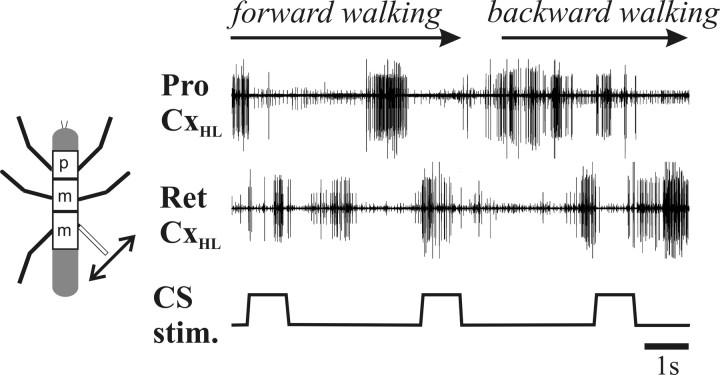
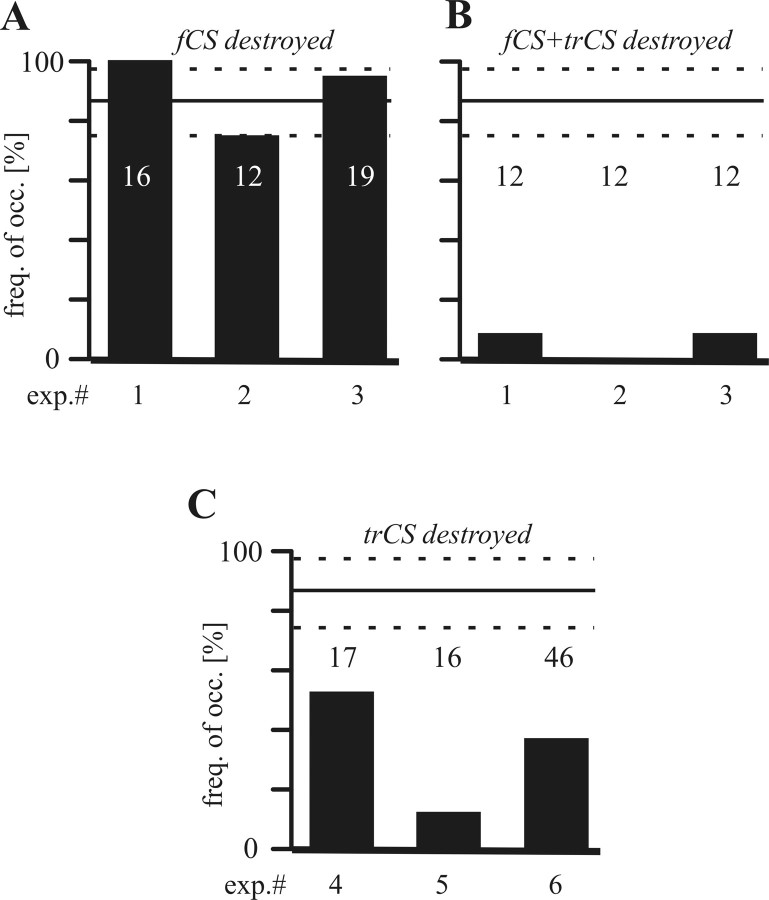
In terrestrial locomotion, sensory feedback from load sensors is important for altering ongoing motor output on a step-by-step basis. We investigated the influence of load signals from the leg on motoneuron pools of the thorax-coxa (ThC) joint in the stick insect walking system. Load sensors were stimulated during rhythmic, alternating activity in protractor coxae (ProCx) and retractor coxae (RetCx) motoneuron pools. Alternating activity in the segment of interest was induced by mechanical stimulation of the animal or pharmacological activation of the isolated thoracic ganglia. Load signals from the legs altered the timing of ThC motoneuron activity by resetting and entraining the activity of the central rhythm generating network of the ThC joint. In the front and middle legs, load signals induced or promoted RetCx activity and decreased or terminated ProCx activity. In the hindleg, reverse transitions were elicited, with increasing load terminating RetCx and initiating ProCx activity. Studies in semi-intact walking animals showed that the effect of load on the ThC-joint motoneurons depended on walking direction, with increased load promoting the functional stance phase motoneuron pool (in forward walking, RetCx activity; in backward walking, ProCx activity). Thus, we show that modifications of sensory feedback in a locomotor system are related to walking direction. In a final set of ablation experiments, we show that the load influence is mediated by the three groups of trochanteral campaniform sensilla.
Figures
References
-
- Akay T, Bässler U, Gerharz P, Büschges A. Interjoint coordination: the role of sensory signals from the coxa-trochanteral joint on controlling motoneuronal activity of the femur-tibia joint in stick insects. J Neurophysiol. 2001;85:594–604. - PubMed
-
- Akay T, Haehn S, Schmitz J, Büschges A. Signals from load sensors underlie interjoint coordination during stepping movements of the stick insect legs. J Neurophysiol. 2004;92:42–51. - PubMed
-
- Ayers JL, Davis WJ. Neuronal control of locomotion in the lobster Homarus americanus. J Comp Physiol. 1977;115:29–46.
-
- Bartling CH, Schmitz J. Reaction to disturbances of a walking leg during stance. J Exp Biol. 2000;203:1211–1233. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources