

Modulation of the cold-activated channel TRPM8 by lysophospholipids and polyunsaturated fatty acids
- PMID: 17376995
- PMCID: PMC2726637
- DOI: 10.1523/JNEUROSCI.4846-06.2007
Modulation of the cold-activated channel TRPM8 by lysophospholipids and polyunsaturated fatty acids
Abstract
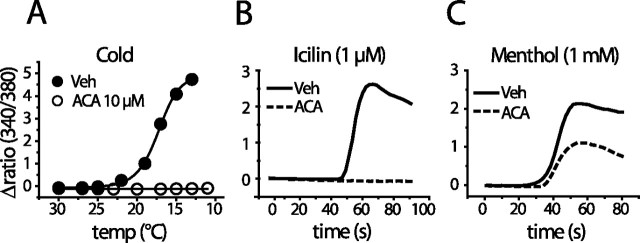
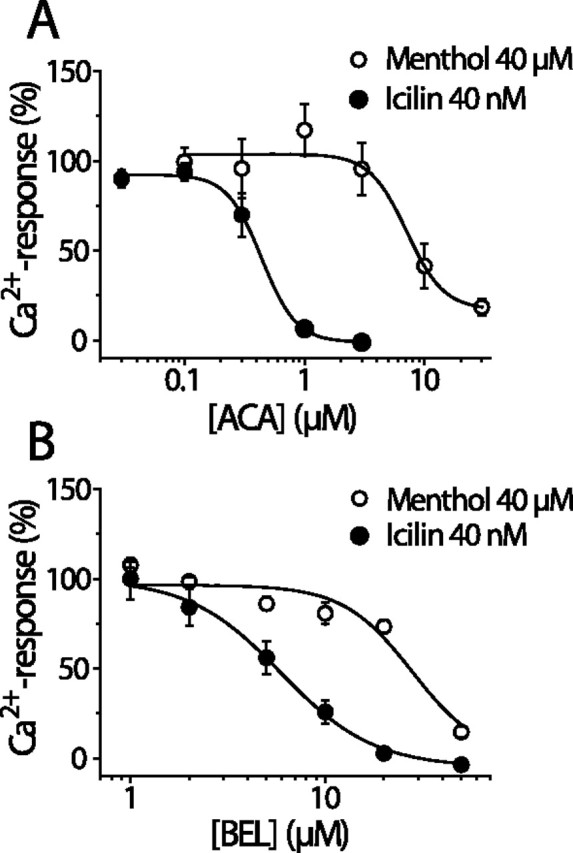
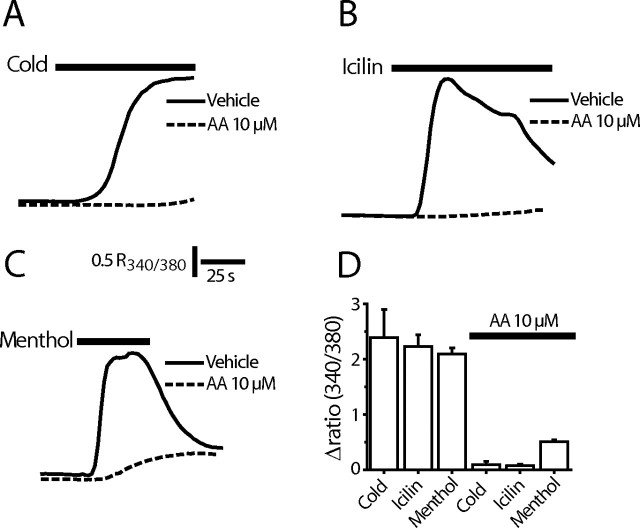
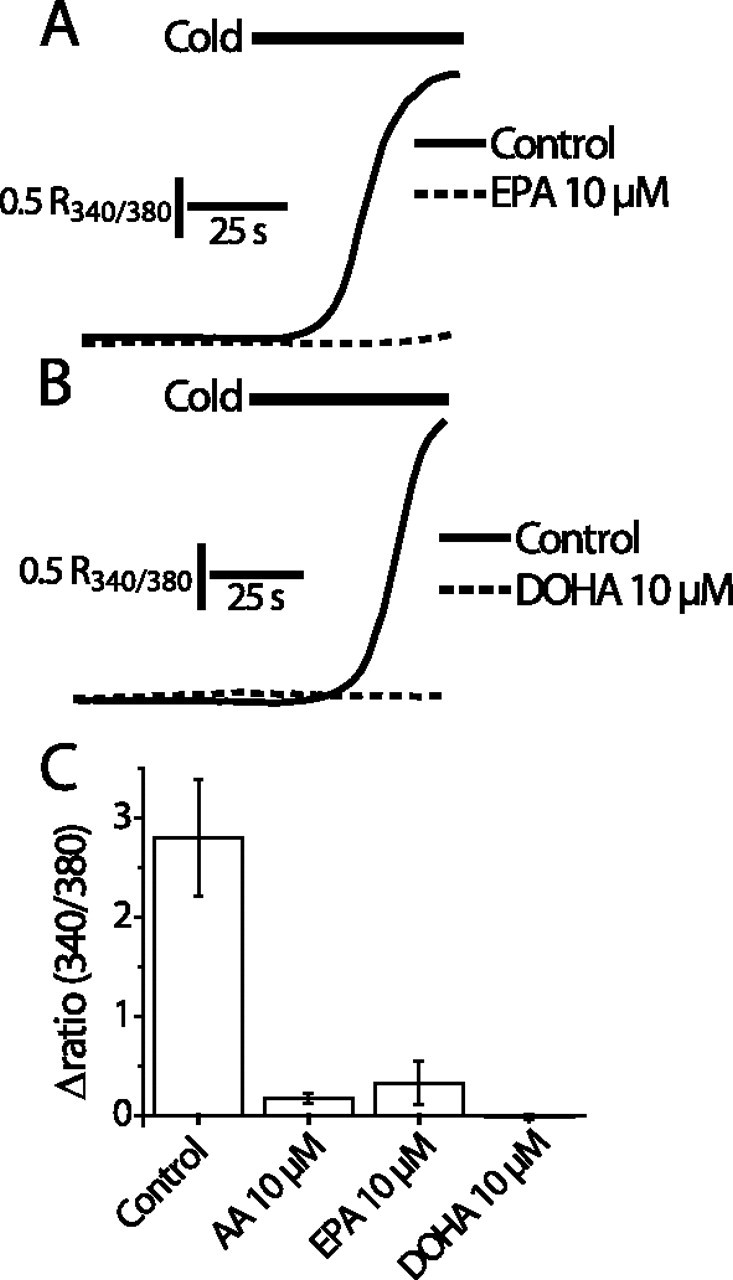
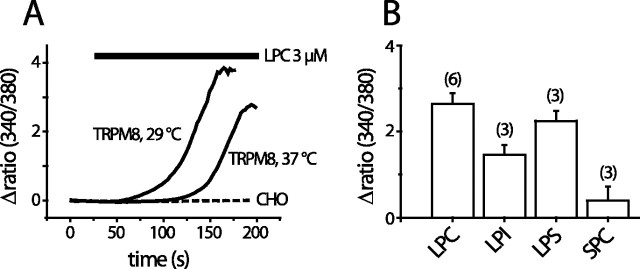
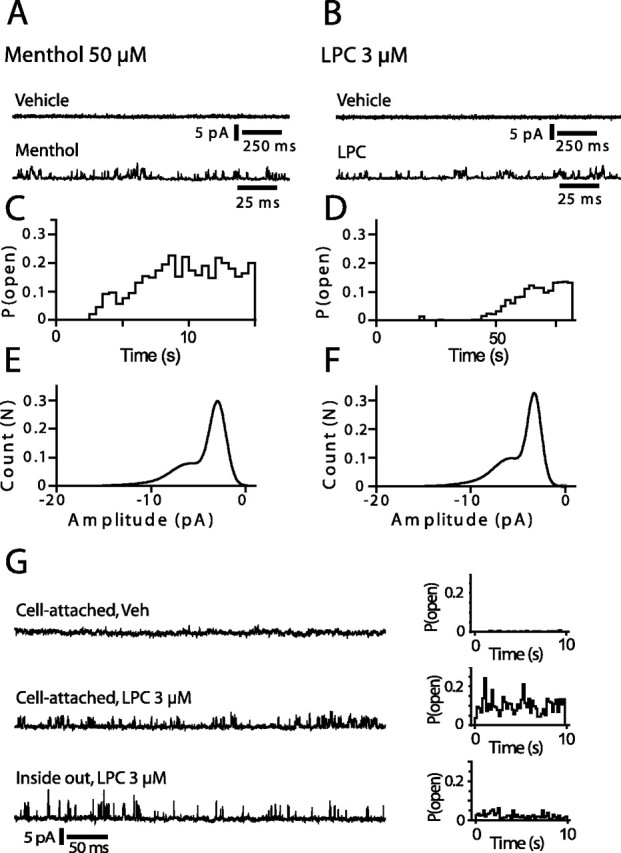
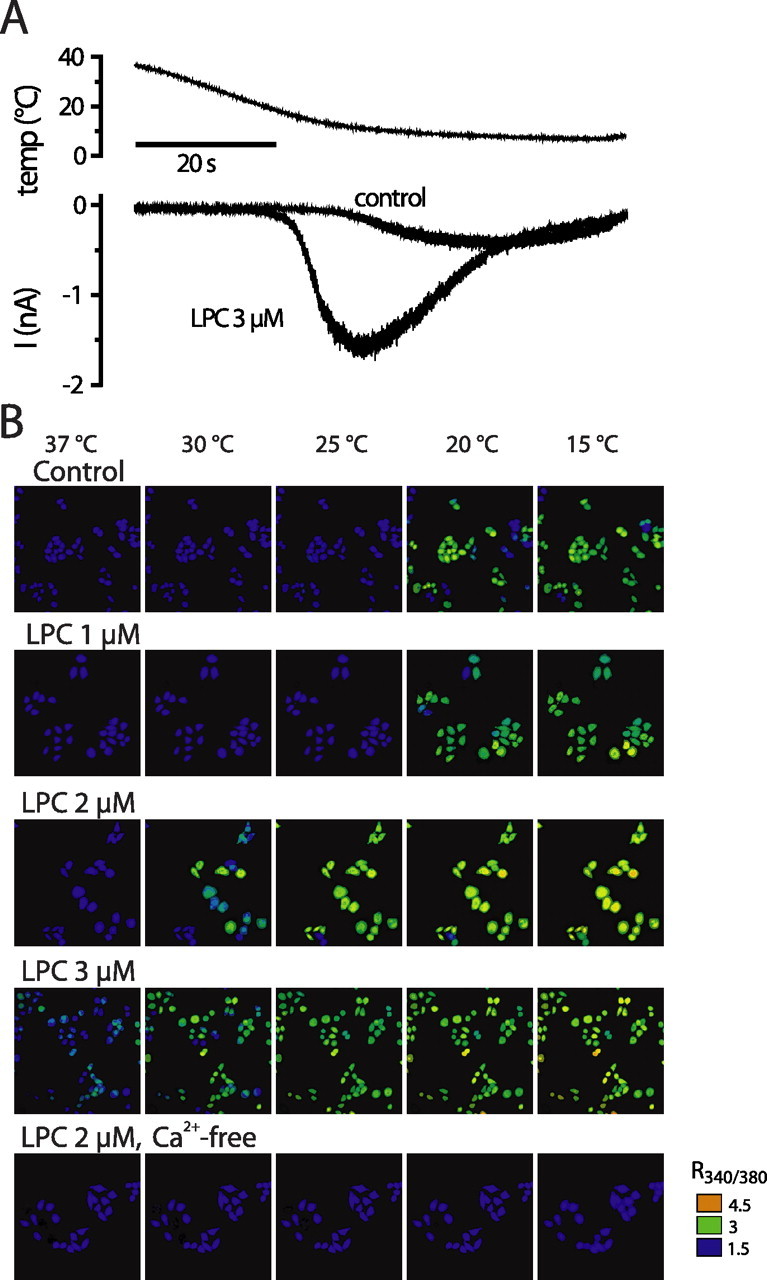
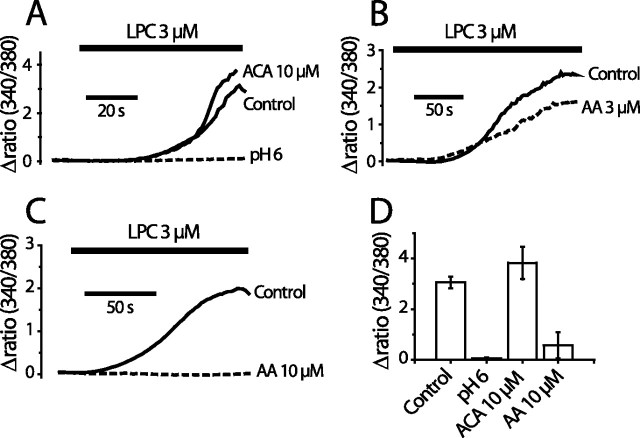
We investigated the role of phospholipase A2 (PLA2) and the effects of PLA2 products (polyunsaturated fatty acids and lysophospholipids) on the cold-sensitive channel transient receptor potential (melastatin)-8 (TRPM8), heterologously expressed in Chinese hamster ovary cells. TRPM8 responses to cold and the agonist icilin were abolished by inhibitors of the calcium-independent (iPLA2) form of the enzyme, whereas responses to menthol were less sensitive to iPLA2 inhibition. Inhibition of PLA2 similarly abolished the cold responses of the majority of cold-sensitive dorsal root ganglion neurons. The products of PLA2 had opposing effects on TRPM8. Lysophospholipids (LPLs) (lysophosphatidylcholine, lysophosphatidylinositol, and lysophosphatidylserine) altered the thermal sensitivity of TRPM8, raising the temperature threshold toward normal body temperature. Polyunsaturated fatty acids (PUFAs), such as arachidonic acid, inhibited the activation of TRPM8 by cold, icilin, and menthol. The relative potencies of lysophospholipids and PUFAs are such that lysophosphatidylcholine is able to modulate TRPM8 in the presence of an equimolar concentration of arachidonic acid. Positive modulation by LPLs provides a potential physiological mechanism for sensitizing and activating TRPM8 in the absence of temperature variations.
Figures
Comment in
-
Warming up the cold reception at a TRPM8 function.J Neurosci. 2007 Jul 18;27(29):7617-8. doi: 10.1523/JNEUROSCI.2353-07.2007. J Neurosci. 2007. PMID: 17634355 Free PMC article. No abstract available.
Similar articles
-
The roles of iPLA2, TRPM8 and TRPA1 in chemically induced cold hypersensitivity.Mol Pain. 2010 Jan 21;6:4. doi: 10.1186/1744-8069-6-4. Mol Pain. 2010. PMID: 20092626 Free PMC article.
-
Camphor activates and sensitizes transient receptor potential melastatin 8 (TRPM8) to cooling and icilin.Chem Senses. 2013 Sep;38(7):563-75. doi: 10.1093/chemse/bjt027. Epub 2013 Jul 4. Chem Senses. 2013. PMID: 23828908
-
Ca2+-independent phospholipase A2-dependent gating of TRPM8 by lysophospholipids.J Biol Chem. 2006 Dec 29;281(52):40174-82. doi: 10.1074/jbc.M605779200. Epub 2006 Nov 2. J Biol Chem. 2006. PMID: 17082190
-
Potential Role of TRPM8 in Cold-Induced Hypertension and Its Clinical Implications.Discov Med. 2023 Aug;35(177):451-457. doi: 10.24976/Discov.Med.202335177.46. Discov Med. 2023. PMID: 37553299 Review.
-
The emerging pharmacology of TRPM8 channels: hidden therapeutic potential underneath a cold surface.Curr Pharm Biotechnol. 2011 Jan 1;12(1):54-67. doi: 10.2174/138920111793937916. Curr Pharm Biotechnol. 2011. PMID: 20932258 Review.
Cited by
-
Scraping through the ice: uncovering the role of TRPM8 in cold transduction.Am J Physiol Regul Integr Comp Physiol. 2011 Jun;300(6):R1278-87. doi: 10.1152/ajpregu.00631.2010. Epub 2011 Mar 16. Am J Physiol Regul Integr Comp Physiol. 2011. PMID: 21411765 Free PMC article. Review.
-
Regulation of thermoTRPs by lipids.Temperature (Austin). 2016 Nov 1;4(1):24-40. doi: 10.1080/23328940.2016.1254136. eCollection 2017. Temperature (Austin). 2016. PMID: 28349093 Free PMC article. Review.
-
TRP (transient receptor potential) ion channel family: structures, biological functions and therapeutic interventions for diseases.Signal Transduct Target Ther. 2023 Jul 5;8(1):261. doi: 10.1038/s41392-023-01464-x. Signal Transduct Target Ther. 2023. PMID: 37402746 Free PMC article. Review.
-
Functional and Modeling Studies of the Transmembrane Region of the TRPM8 Channel.Biophys J. 2015 Nov 3;109(9):1840-51. doi: 10.1016/j.bpj.2015.09.027. Biophys J. 2015. PMID: 26536261 Free PMC article.
-
Lysophospholipids Contribute to Oxaliplatin-Induced Acute Peripheral Pain.J Neurosci. 2020 Dec 2;40(49):9519-9532. doi: 10.1523/JNEUROSCI.1223-20.2020. Epub 2020 Nov 6. J Neurosci. 2020. PMID: 33158961 Free PMC article.
References
-
- Abeele FV, Zholos A, Bidaux G, Shuba Y, Thebault S, Beck B, Flourakis M, Panchin Y, Skryma R, Prevarskaya N. Ca(2+)-independent phospholipase A(2)-dependent gating of TRPM8 by lysophospholipids. J Biol Chem. 2006;281:40174–40182. - PubMed
-
- Babes A, Zorzon D, Reid G. Two populations of cold-sensitive neurons in rat dorsal root ganglia and their modulation by nerve growth factor. Eur J Neurosci. 2004;20:2276–2282. - PubMed
-
- Balsinde J, Balboa MA. Cellular regulation and proposed biological functions of group VIA calcium-independent phospholipase A2 in activated cells. Cell Signal. 2005;17:1052–1062. - PubMed
-
- Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron. 2004;41:849–857. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources