

The role of 3-canal biomechanics in angular motion transduction by the human vestibular labyrinth
- PMID: 17377842
- PMCID: PMC3005417
- DOI: 10.1007/s10439-007-9277-y
The role of 3-canal biomechanics in angular motion transduction by the human vestibular labyrinth
Abstract
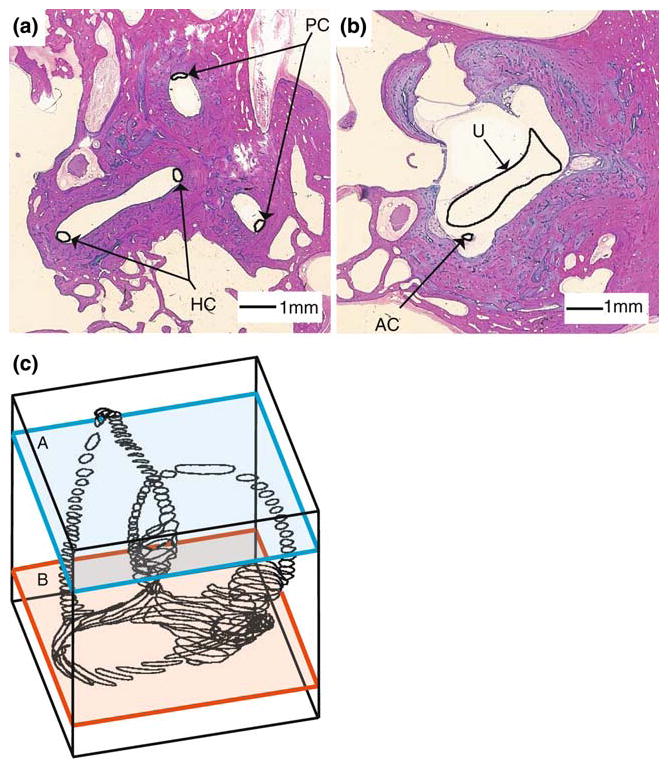
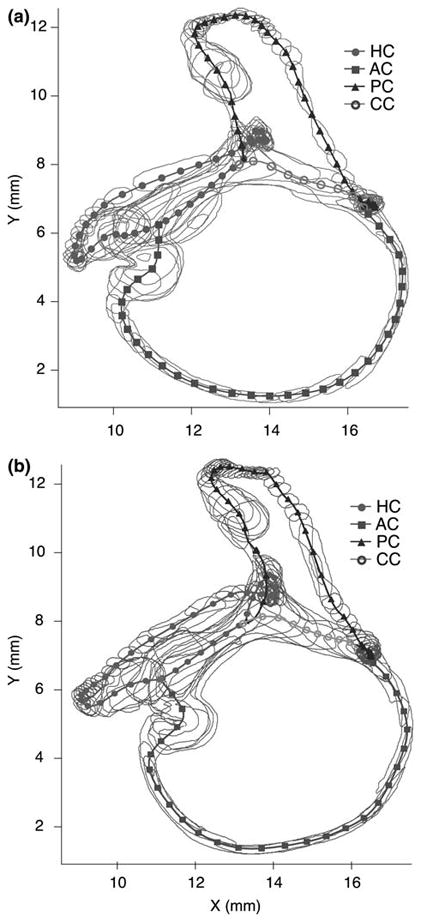
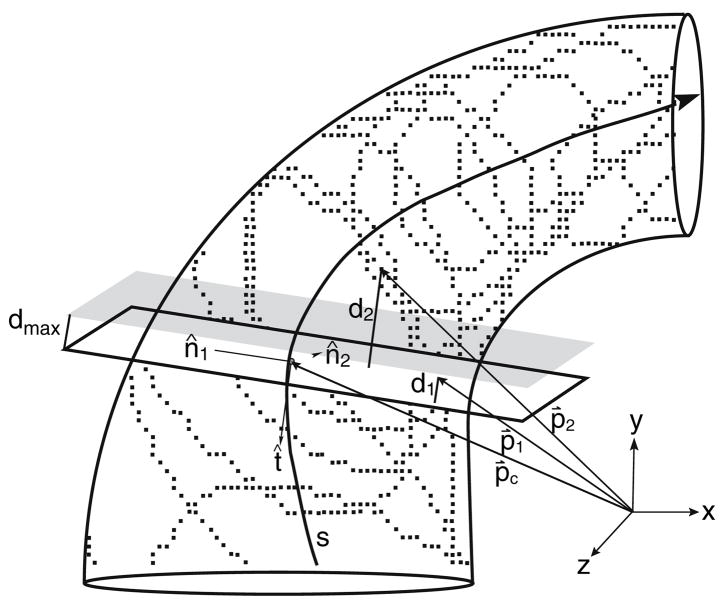
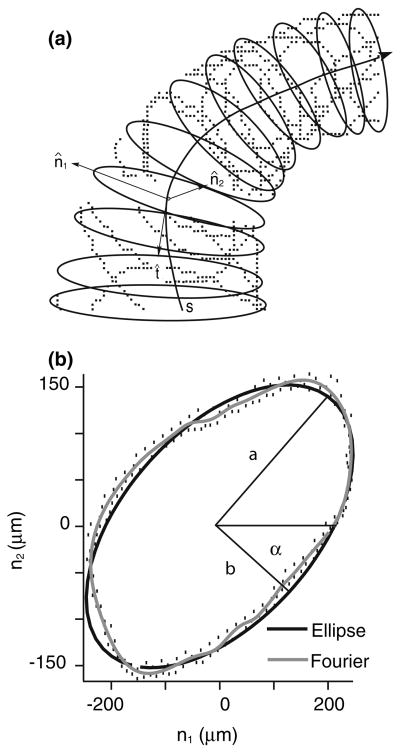
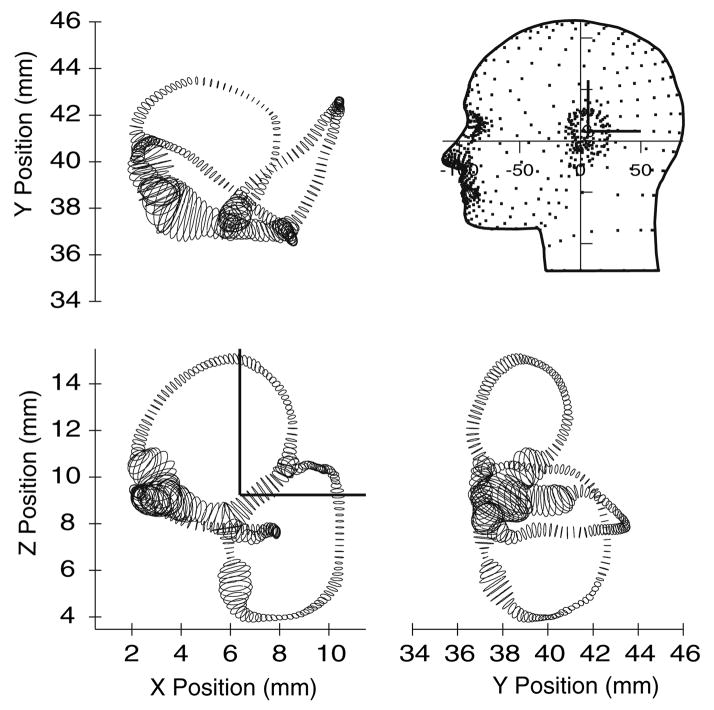
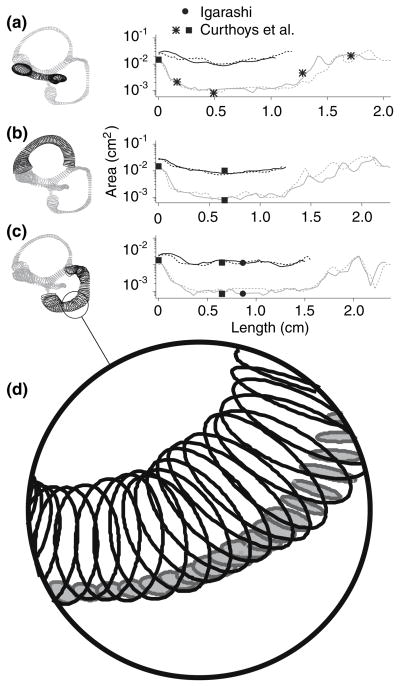
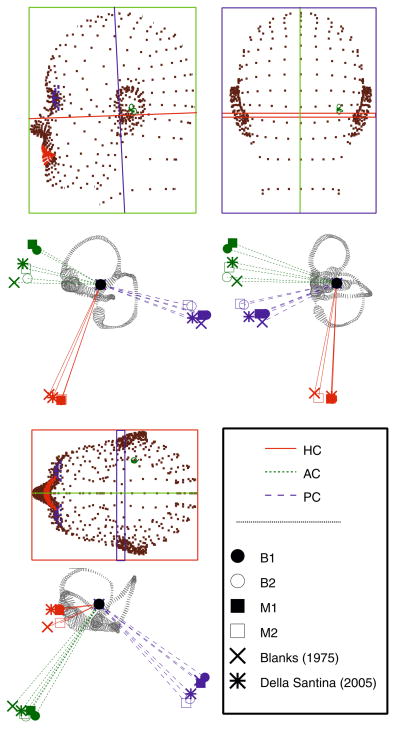
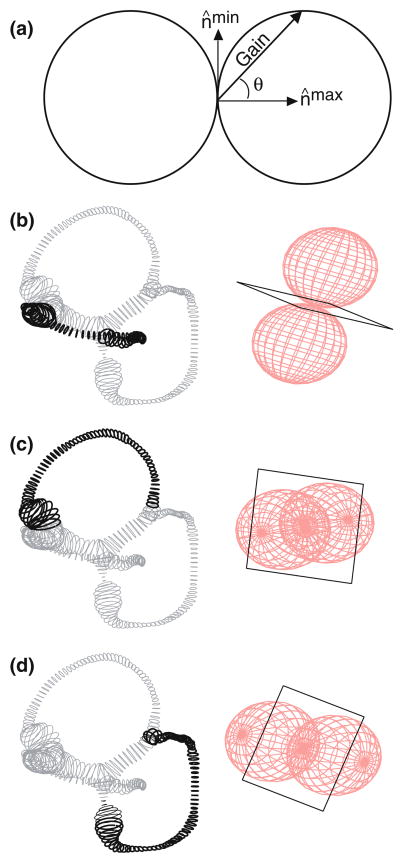
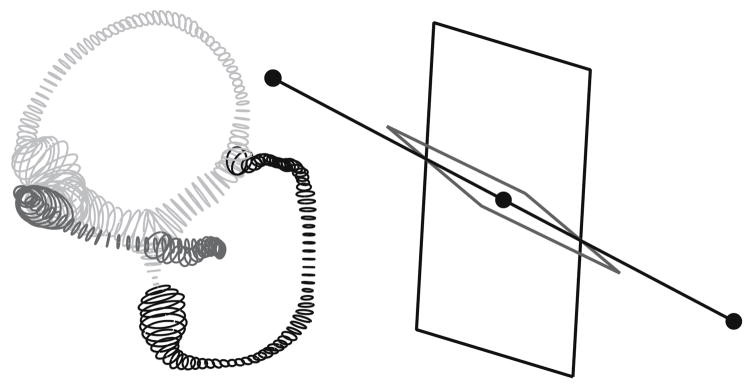
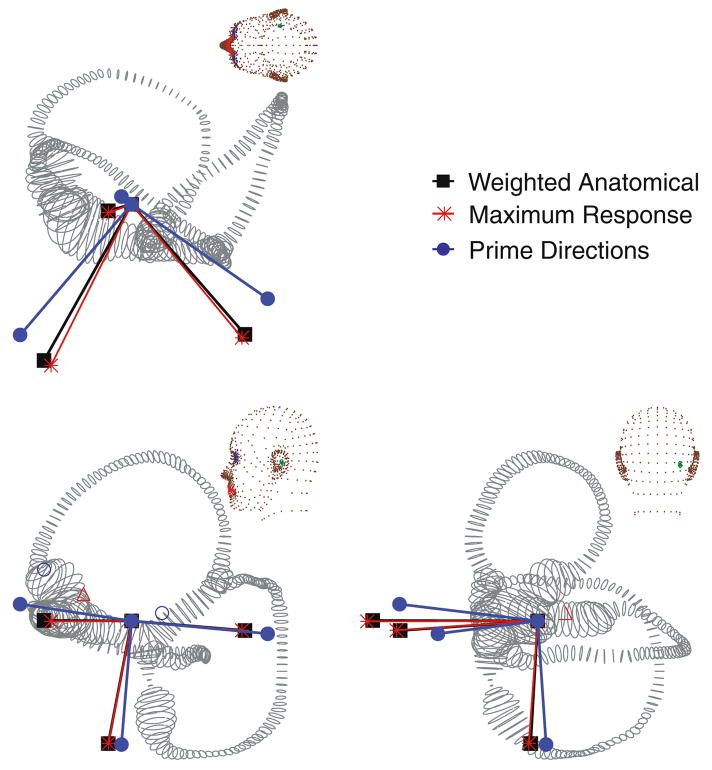
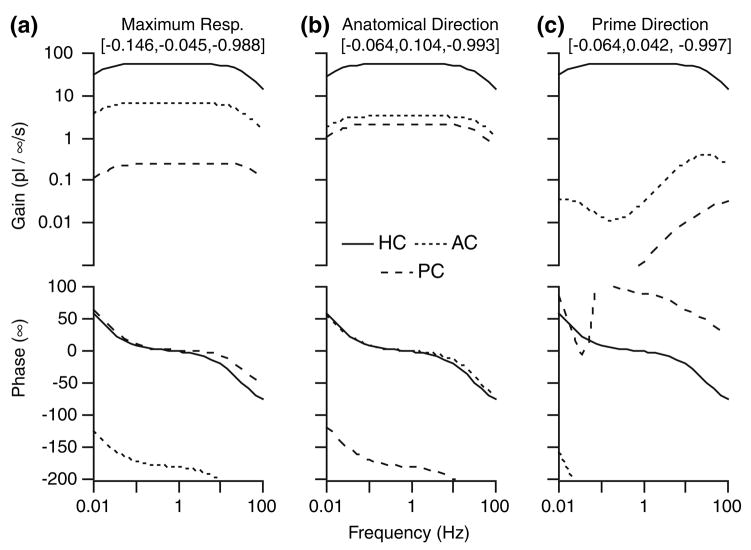
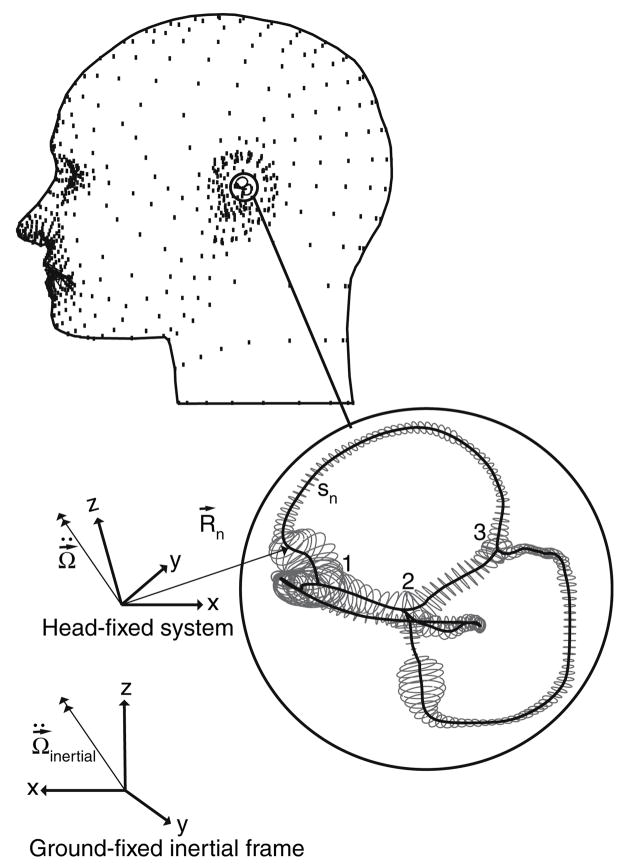
The present work examines the role of the complex geometry of the human vestibular membranous labyrinth in the process of angular motion transduction by the semicircular canals. A morphologically descriptive mathematical model was constructed to address the biomechanical origins of temporal signal processing and directional coding in determining the inputs to the brain. The geometrical model was developed based on shrinkage-corrected temporal bone sections using a segmentation/data-fitting procedure. Endolymph fluid dynamics within the 3-canal labyrinth was modeled using an asymptotic form of the Navier-Stokes equations and solved to estimate endolymph and cupulae volume displacements. The geometrical model was manipulated to study the role of major morphological features on directional and temporal coding. Anatomical results show that the bony osseous canals provide reasonable estimates of the orientation of the delicate membranous canals--the two differed by only 3.48 +/- 1.89 degrees . Biomechanical results show that the maximal response directions are distinct from the anatomical canal planes, but can be closely approximated by fitting a flat plane to the centerline of the canal of interest and weighting each location along the centerline with the inverse of the cross-sectional area squared. Vector cross-products of these maximal response directions, in turn, determine the null planes and prime directions that transmit the direction of angular motion to the brain as three independent directional channels associated with the nerve bundles. Finally, parameter studies indicate that changes in canal cross-sectional area and shape only moderately affect canal temporal and directional coding, while three-canal orientation is critical to directional coding.
Figures
Similar articles
-
Wave Mechanics of the Vestibular Semicircular Canals.Biophys J. 2017 Sep 5;113(5):1133-1149. doi: 10.1016/j.bpj.2017.08.001. Biophys J. 2017. PMID: 28877495 Free PMC article.
-
Directional coding of three-dimensional movements by the vestibular semicircular canals.Biol Cybern. 1999 Jun;80(6):417-31. doi: 10.1007/s004220050536. Biol Cybern. 1999. PMID: 10420568
-
Determinants of spatial and temporal coding by semicircular canal afferents.J Neurophysiol. 2005 May;93(5):2359-70. doi: 10.1152/jn.00533.2004. J Neurophysiol. 2005. PMID: 15845995 Free PMC article. Review.
-
Analysis of Vestibular Labyrinthine Geometry and Variation in the Human Temporal Bone.Front Neurosci. 2018 Feb 26;12:107. doi: 10.3389/fnins.2018.00107. eCollection 2018. Front Neurosci. 2018. PMID: 29535601 Free PMC article.
-
Semicircular canal biomechanics in health and disease.J Neurophysiol. 2019 Mar 1;121(3):732-755. doi: 10.1152/jn.00708.2018. Epub 2018 Dec 19. J Neurophysiol. 2019. PMID: 30565972 Free PMC article. Review.
Cited by
-
BPPV Viewer: A downloadable 3D BPPV model for study of otolith disease.World J Otorhinolaryngol Head Neck Surg. 2020 Jun 8;7(1):34-39. doi: 10.1016/j.wjorl.2018.10.001. eCollection 2021 Jan. World J Otorhinolaryngol Head Neck Surg. 2020. PMID: 33474542 Free PMC article.
-
A mathematical model of human semicircular canal geometry: a new basis for interpreting vestibular physiology.J Assoc Res Otolaryngol. 2010 Jun;11(2):145-59. doi: 10.1007/s10162-009-0195-6. Epub 2009 Dec 1. J Assoc Res Otolaryngol. 2010. PMID: 19949828 Free PMC article.
-
Mechanical amplification by hair cells in the semicircular canals.Proc Natl Acad Sci U S A. 2010 Feb 23;107(8):3864-9. doi: 10.1073/pnas.0906765107. Epub 2010 Feb 4. Proc Natl Acad Sci U S A. 2010. PMID: 20133682 Free PMC article.
-
Human yaw rotation aftereffects with brief duration rotations are inconsistent with velocity storage.J Assoc Res Otolaryngol. 2014 Apr;15(2):305-17. doi: 10.1007/s10162-013-0438-4. Epub 2014 Jan 10. J Assoc Res Otolaryngol. 2014. PMID: 24408345 Free PMC article.
-
Wave Mechanics of the Vestibular Semicircular Canals.Biophys J. 2017 Sep 5;113(5):1133-1149. doi: 10.1016/j.bpj.2017.08.001. Biophys J. 2017. PMID: 28877495 Free PMC article.
References
-
- Arnold B, Jager L, Grevers G. Visualization of inner ear structures by three-dimensional high-resolution magnetic resonance imaging. Am J Otol. 1996;17(3):480–485. - PubMed
-
- Baird RA, et al. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J Neurophysiol. 1988;60(1):182–203. - PubMed
-
- Blanks RH, et al. Planar relationships of the semicircular canals in rhesus and squirrel monkeys. Brain Res. 1985;340(2):315–324. - PubMed
-
- Blanks RH, I, Curthoys S, Markham CH. Planar relationships of semicircular canals in the cat. Am J Physiol. 1972;223(1):55–62. - PubMed
-
- Blanks RH, I, Curthoys S, Markham CH. Planar relationships of the semicircular canals in man. Acta Otolaryngol. 1975;80(3–4):185–196. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
