

Role of lysine187 within the second extracellular loop of the type A cholecystokinin receptor in agonist-induced activation. Use of complementary charge-reversal mutagenesis to define a functionally important interdomain interaction
- PMID: 17381074
- PMCID: PMC2580722
- DOI: 10.1021/bi0622468
Role of lysine187 within the second extracellular loop of the type A cholecystokinin receptor in agonist-induced activation. Use of complementary charge-reversal mutagenesis to define a functionally important interdomain interaction
Abstract
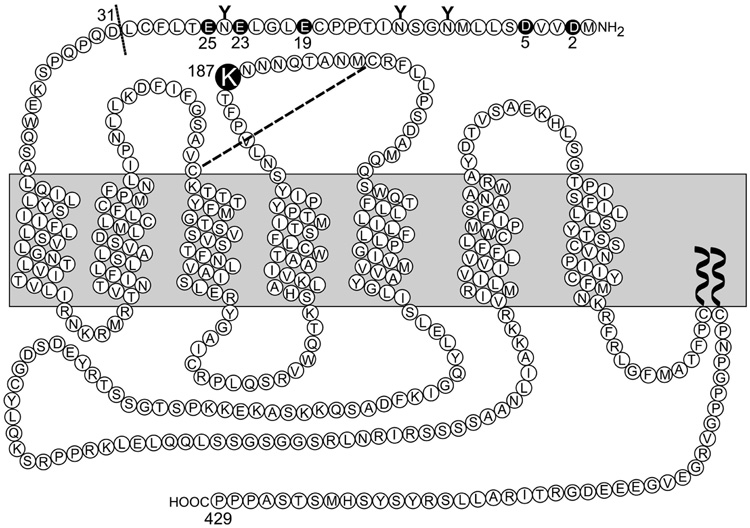
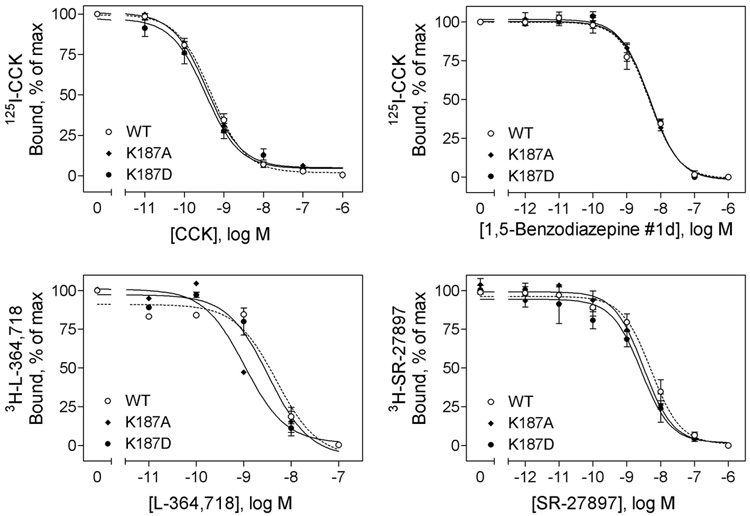
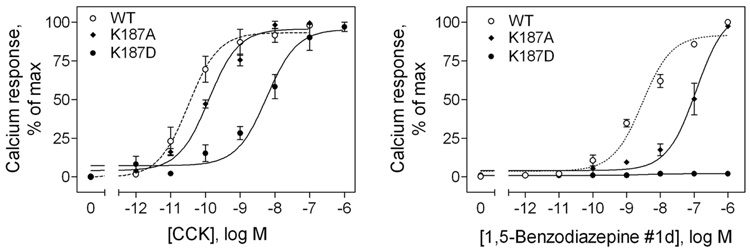
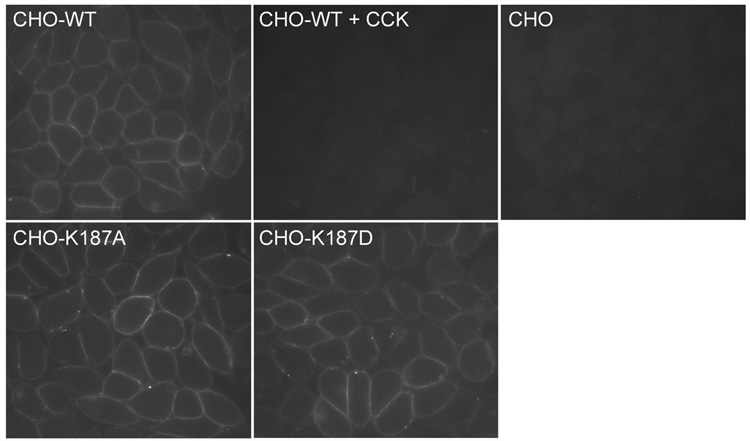
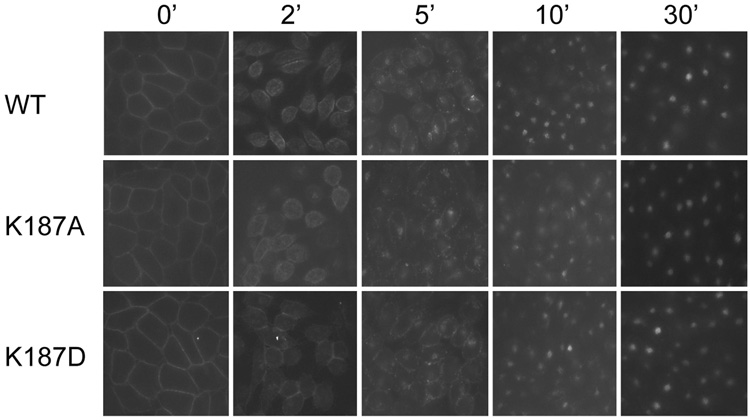
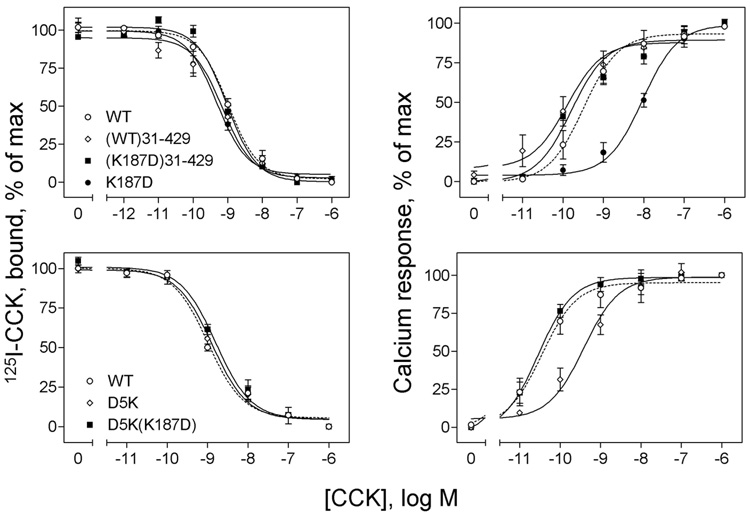
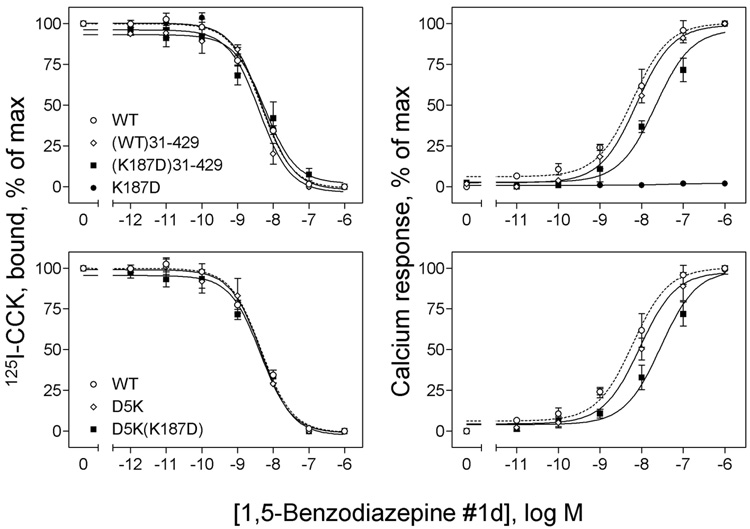
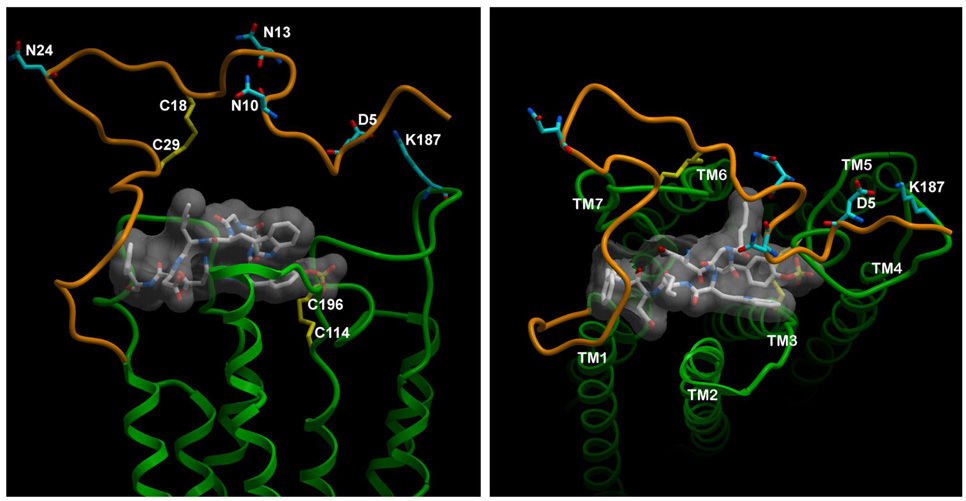
Activation of guanine nucleotide-binding protein (G protein)-coupled receptors is believed to involve conformational change that exposes a domain for G protein coupling at the cytosolic surface of the helical confluence, although the mechanisms for achieving this are not well understood. This conformational change can be achieved by docking a diverse variety of agonist ligands, known to occur by interacting with different regions of these receptors. In this study, we focus on the importance of a specific basic residue (Lys187) within the second extracellular loop of the receptor for the peptide hormone, cholecystokinin. Alanine-replacement and charge-reversal mutagenesis of this residue showed that it had no effect on the binding of natural peptide and nonpeptidyl ligands of this receptor but markedly interfered with agonist-stimulated signaling. It was demonstrated that this negative effect on biological activity could be eliminated with the truncation of the first 30 residues of the amino-terminal tail of this receptor. Complementary charge-reversal mutagenesis of each of the five conserved acidic residues within this region of the receptor in the presence of the charge-reversed Lys187 revealed that only the Asp5 mutant fully reversed the negative functional impact of the Lys187 charge reversal. Thus, we have demonstrated that a basic residue within the second extracellular loop of the cholecystokinin receptor interacts with a specific acidic residue within the amino terminus of this receptor. This residue-residue interaction is nicely accommodated within a new molecular model of the agonist-occupied cholecystokinin receptor.
Figures
Similar articles
-
Elucidation of the molecular basis of cholecystokinin Peptide docking to its receptor using site-specific intrinsic photoaffinity labeling and molecular modeling.Biochemistry. 2009 Jun 16;48(23):5303-12. doi: 10.1021/bi9004705. Biochemistry. 2009. PMID: 19441839 Free PMC article.
-
Refinement of the conformation of a critical region of charge-charge interaction between cholecystokinin and its receptor.Mol Pharmacol. 2002 May;61(5):1041-52. doi: 10.1124/mol.61.5.1041. Mol Pharmacol. 2002. PMID: 11961122
-
Molecular basis of agonist binding to the type A cholecystokinin receptor.Pharmacol Toxicol. 2002 Dec;91(6):282-5. doi: 10.1034/j.1600-0773.2002.910603.x. Pharmacol Toxicol. 2002. PMID: 12688369 Review.
-
Measurement of intermolecular distances for the natural agonist Peptide docked at the cholecystokinin receptor expressed in situ using fluorescence resonance energy transfer.Mol Pharmacol. 2004 Jan;65(1):28-35. doi: 10.1124/mol.65.1.28. Mol Pharmacol. 2004. PMID: 14722234
-
Characterization of the type A cholecystokinin receptor hormone-binding domain: use of contrasting and complementary methodologies.Peptides. 2001 Aug;22(8):1223-8. doi: 10.1016/s0196-9781(01)00445-4. Peptides. 2001. PMID: 11457514 Review.
Cited by
-
Elucidation of the molecular basis of cholecystokinin Peptide docking to its receptor using site-specific intrinsic photoaffinity labeling and molecular modeling.Biochemistry. 2009 Jun 16;48(23):5303-12. doi: 10.1021/bi9004705. Biochemistry. 2009. PMID: 19441839 Free PMC article.
-
The micelle-associated 3D structures of Boc-Y(SO3)-Nle-G-W-Nle-D-2-phenylethylester (JMV-180) and CCK-8(s) share conformational elements of a calculated CCK1 receptor-bound model.J Med Chem. 2008 Jul 10;51(13):3742-54. doi: 10.1021/jm701401j. Epub 2008 Jun 10. J Med Chem. 2008. PMID: 18540665 Free PMC article.
-
Development of a highly selective allosteric antagonist radioligand for the type 1 cholecystokinin receptor and elucidation of its molecular basis of binding.Mol Pharmacol. 2015 Jan;87(1):130-40. doi: 10.1124/mol.114.095430. Epub 2014 Oct 15. Mol Pharmacol. 2015. PMID: 25319540 Free PMC article.
-
Molecular basis for agonist selectivity and activation of the orphan bombesin receptor subtype 3 receptor.J Pharmacol Exp Ther. 2008 Feb;324(2):463-74. doi: 10.1124/jpet.107.132332. Epub 2007 Nov 15. J Pharmacol Exp Ther. 2008. PMID: 18006692 Free PMC article.
-
Benzodiazepine ligands can act as allosteric modulators of the Type 1 cholecystokinin receptor.Bioorg Med Chem Lett. 2008 Aug 1;18(15):4401-4. doi: 10.1016/j.bmcl.2008.06.053. Epub 2008 Jun 20. Bioorg Med Chem Lett. 2008. PMID: 18621527 Free PMC article.
References
-
- Oliveira L, Paiva PB, Paiva AC, Vriend G. Sequence analysis reveals how G protein-coupled receptors transduce the signal to the G protein. Proteins. 2003;52:553–560. - PubMed
-
- Oliveira L, Paiva AC, Vriend G. Correlated mutation analyses on very large sequence families. Chembiochem. 2002;3:1010–1017. - PubMed
-
- Schmitz F, Pratt DS, Wu MJ, Kolakowski LF, Jr, Beinborn M, Kopin AS. Identification of cholecystokinin-B/gastrin receptor domains that confer high gastrin affinity: utilization of a novel Xenopus laevis cholecystokinin receptor. Mol Pharmacol. 1996;50:436–441. - PubMed
-
- Noble F, Wank SA, Crawley JN, Bradwejn J, Seroogy KB, Hamon M, Roques BP. International Union of Pharmacology. XXI. Structure, distribution, and functions of cholecystokinin receptors. Pharmacol Rev. 1999;51:745–781. - PubMed
-
- Ding XQ, Pinon DI, Furse KE, Lybrand TP, Miller LJ. Refinement of the conformation of a critical region of charge-charge interaction between cholecystokinin and its receptor. Mol Pharmacol. 2002;61:1041–1052. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
