

Synergistic substrate inhibition of ent-copalyl diphosphate synthase: a potential feed-forward inhibition mechanism limiting gibberellin metabolism
- PMID: 17384166
- PMCID: PMC1913771
- DOI: 10.1104/pp.106.095208
Synergistic substrate inhibition of ent-copalyl diphosphate synthase: a potential feed-forward inhibition mechanism limiting gibberellin metabolism
Abstract
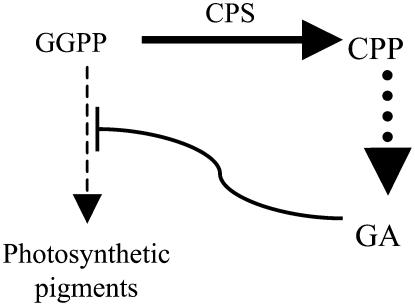
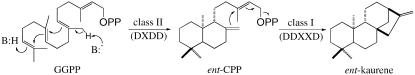
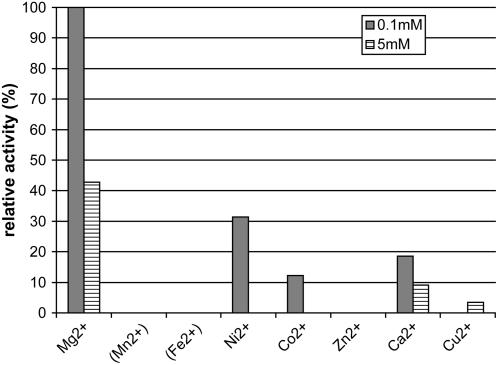
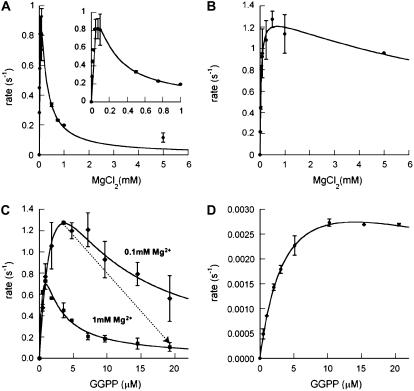
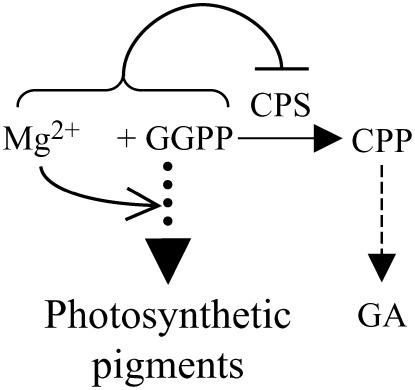
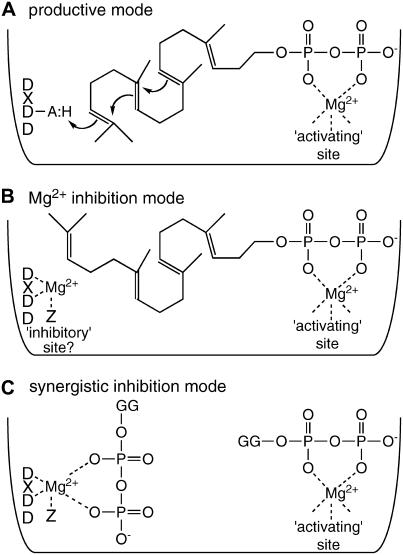
Gibberellins (GAs) or gibberellic acids are ubiquitous diterpenoid phytohormones required for many aspects of plant growth and development, including repression of photosynthetic pigment production (i.e. deetiolation) in the absence of light. The committed step in GA biosynthesis is catalyzed in plastids by ent-copalyl diphosphate synthase (CPS), whose substrate, (E,E,E,)-geranylgeranyl diphosphate (GGPP), is also a direct precursor of carotenoids and the phytol side chain of chlorophyll. Accordingly, during deetiolation, GA production is repressed, whereas flux toward these photosynthetic pigments through their common GGPP precursor is dramatically increased. How this is accomplished has been unclear because no mechanism for regulation of CPS activity has been reported. We present here kinetic analysis of recombinant pseudomature CPS from Arabidopsis (Arabidopsis thaliana; rAtCPS) demonstrating that Mg(2+) and GGPP exert synergistic substrate inhibition effects on CPS activity. These results suggest that GA metabolism may be limited by feed-forward inhibition of CPS; in particular, the effect of Mg(2+) because light induces increases in plastid Mg(2+) levels over a similar range as that observed here to affect rAtCPS activity. Notably, this effect is most pronounced in the GA-specific AtCPS because the corresponding activity of the resin acid biosynthetic enzyme abietadiene synthase is 100-fold less sensitive to [Mg(2+)]. Furthermore, Mg(2+) allosterically activates the plant porphobilinogen synthase involved in chlorophyll production. Hence, Mg(2+) may have a broad role in regulating plastidial metabolic flux during deetiolation. Finally, the observed synergistic substrate/feed-forward inhibition of CPS also seems to provide a novel example of direct regulation of enzymatic activity in hormone biosynthesis.
Figures
References
-
- Aach H, Bose G, Graebe JE (1995) ent-Kaurene biosynthesis in a cell-free system from wheat (Triticum aestivum L.) seedlings and the localisation of ent-kaurene synthetase in plastids of three species. Planta 197 333–342
-
- Ait-Ali T, Swain SM, Reid JB, Sun T, Kamiya Y (1997) The LS locus of pea encodes the gibberellin biosynthesis enzyme ent-kaurene synthase A. Plant J 11 443–454 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
