

Bub1 is essential for assembly of the functional inner centromere
- PMID: 17389228
- PMCID: PMC2064078
- DOI: 10.1083/jcb.200609044
Bub1 is essential for assembly of the functional inner centromere
Abstract
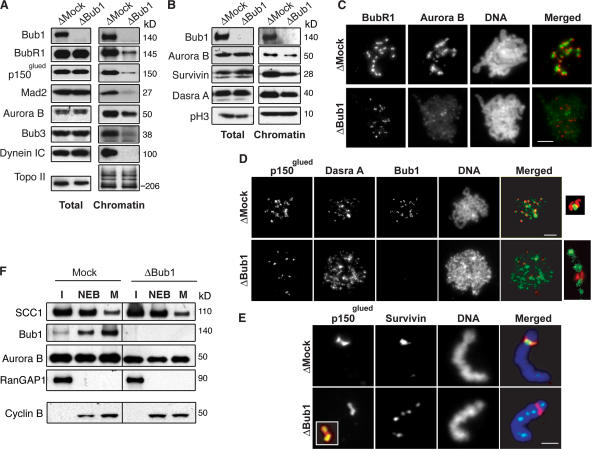
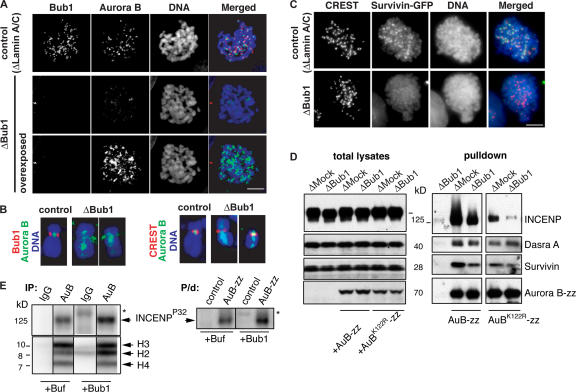
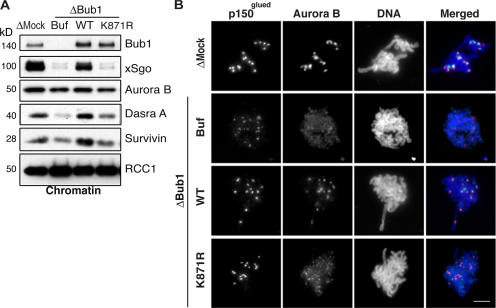
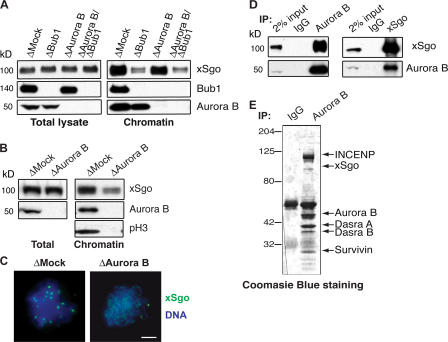
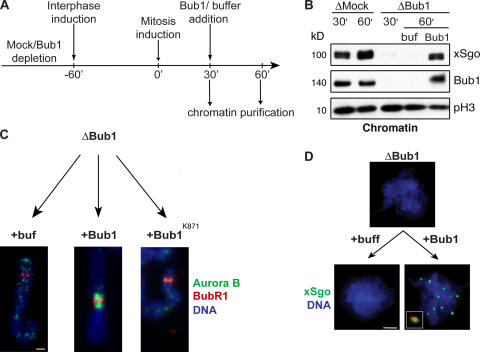
During mitosis, the inner centromeric region (ICR) recruits protein complexes that regulate sister chromatid cohesion, monitor tension, and modulate microtubule attachment. Biochemical pathways that govern formation of the inner centromere remain elusive. The kinetochore protein Bub1 was shown to promote assembly of the outer kinetochore components, such as BubR1 and CENP-F, on centromeres. Bub1 was also implicated in targeting of Shugoshin (Sgo) to the ICR. We show that Bub1 works as a master organizer of the ICR. Depletion of Bub1 from Xenopus laevis egg extract or from HeLa cells resulted in both destabilization and displacement of chromosomal passenger complex (CPC) from the ICR. Moreover, soluble Bub1 controls the binding of Sgo to chromatin, whereas the CPC restricts loading of Sgo specifically onto centromeres. We further provide evidence that Bub1 kinase activity is pivotal for recruitment of all of these components. Together, our findings demonstrate that Bub1 acts at multiple points to assure the correct kinetochore formation.
Figures

References
-
- Andrews, P.D., E. Knatko, W.J. Moore, and J.R. Swedlow. 2003. Mitotic mechanics: the auroras come into view. Curr. Opin. Cell Biol. 15:672–683. - PubMed
-
- Andrews, P.D., Y. Ovechkina, N. Morrice, M. Wagenbach, K. Duncan, L. Wordeman, and J.R. Swedlow. 2004. Aurora B regulates MCAK at the mitotic centromere. Dev. Cell. 6:253–268. - PubMed
-
- Chan, G.K., S.T. Liu, and T.J. Yen. 2005. Kinetochore structure and function. Trends Cell Biol. 15:589–598. - PubMed
